参考资料:《Cambrian spiral-plated echinoderms from Gondwana reveal the earliest pentaradial body plan》(Andrew B. Smith、 Samuel Zamora)
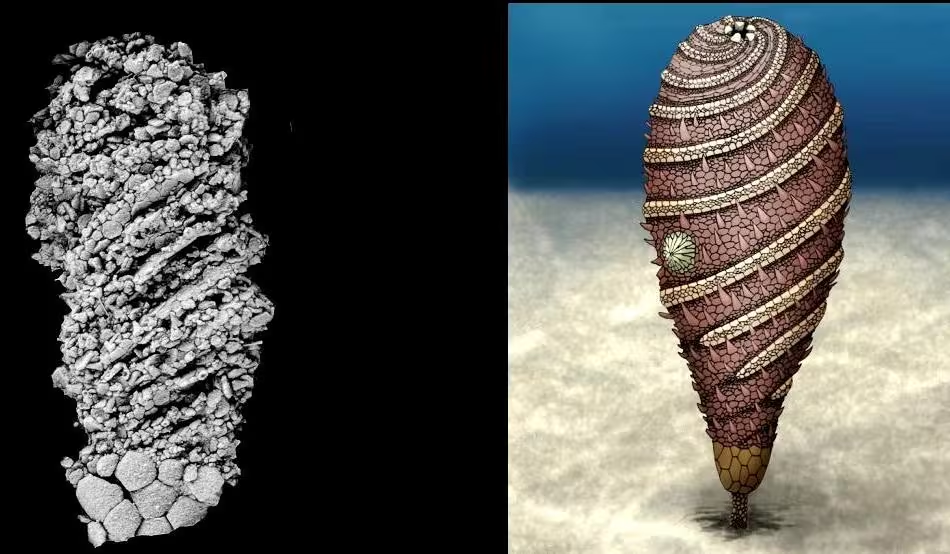
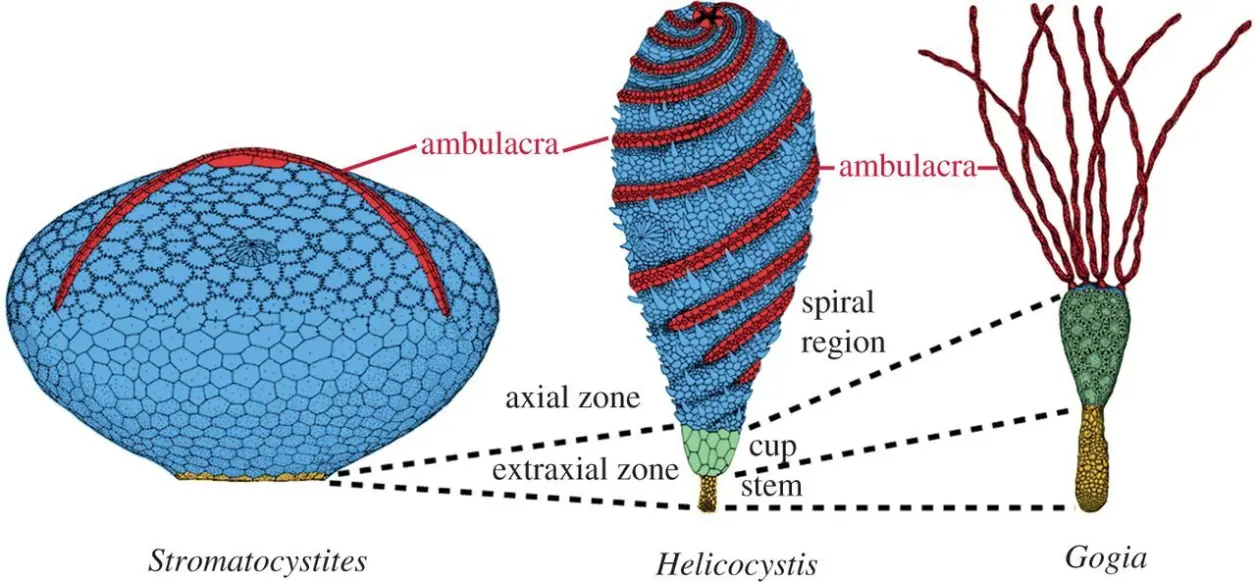
形态描述:本种全长(高)5到20毫米(不计末端固着器),躯体被纵向划分为茎、萼、螺旋域三段,可能分别由幼虫的轴腔、水腔和后体腔发育而来。亚成体的螺旋域与萼部大小相仿,但前者的体积占比会在生长过程中不断增大。螺旋域的外表面覆盖着多列螺旋排布的骨板,分别构成了五条步带和五条间步带。其中,间步带由一系列多边形骨板(越靠近两侧的步带,骨板越小)镶嵌而成,其狭窄的铰接面具有较深的凹槽,使这一区域可能具有手凤琴音箱式的伸缩能力;而步带则从围口域螺旋式地放射出来,直至终止于螺旋域的下端(永远不会触及萼部)。两列缺乏穿孔的底板(florring plate)托举着每条步带的食物沟,具短突的初级盖板(primary covering plate)则与底板一一配对,步带中线的两边还有次级盖板(secondary cover plates),但由于过于微小,很难被完整地保存下来。口部则位于螺旋域的顶部,其周边排布着七片环口骨板:若将C/D间步带上的三片视作一片①(此三者构成了水孔/生殖孔的边界,),则这“五片”环口骨板的布局已经相当接近典型的“2-1-2”模式②。围肛部位于螺旋域的侧面中部。螺旋域的下方,与之界限分明的第二个体段——萼部,则覆盖着两到三圈粗大而厚实的多边形骨片:它们彼此紧密拼合,使萼部具有了坚固的杯状框架。最后,在萼部逐渐收窄的底部,出现了短而细的茎(stem)或梗(peduncle),该体段覆盖着小而无序的圆形骨片,它们既不镶嵌,也不叠覆,而仅仅是松散地毗邻着。
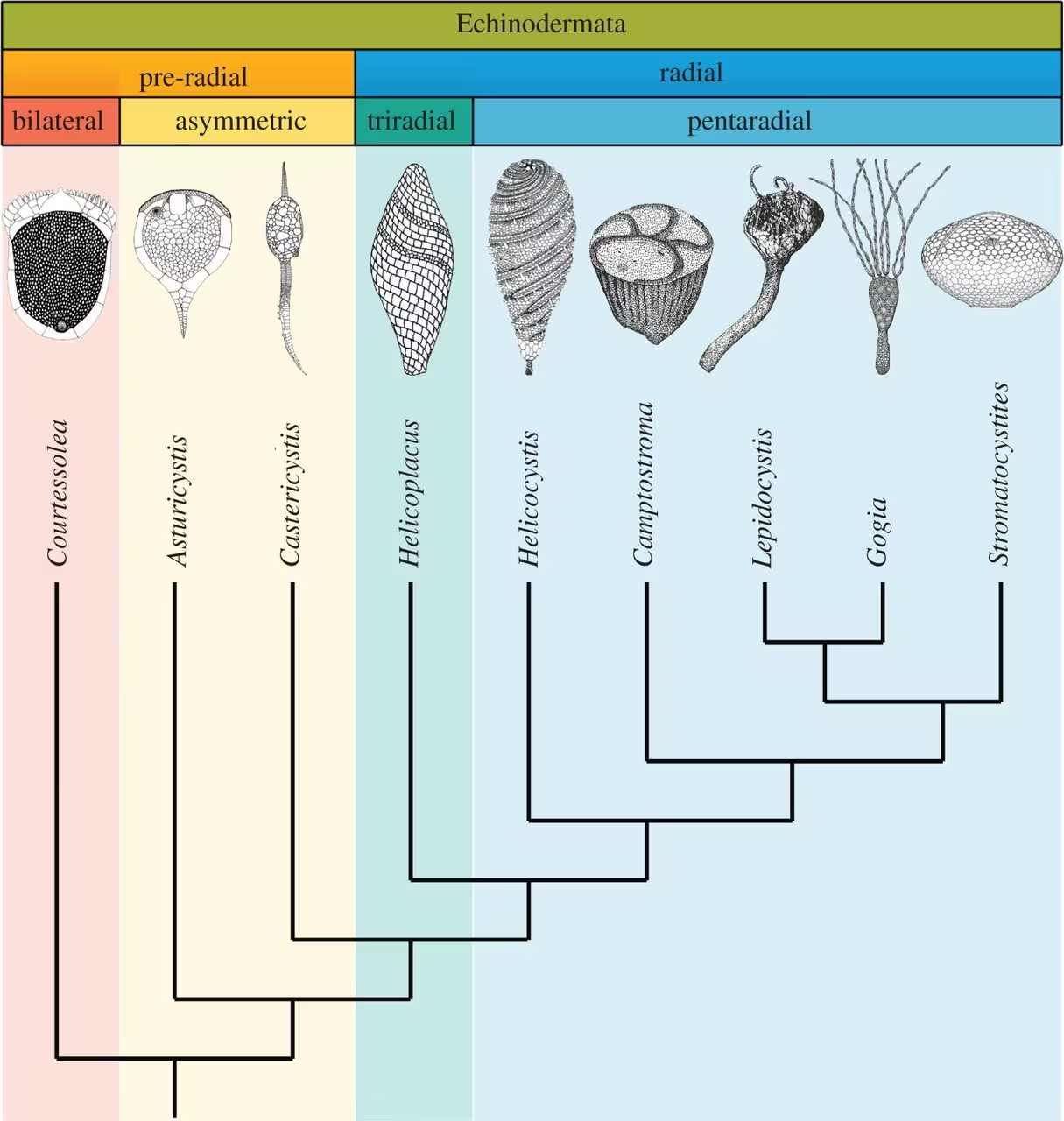
演化意义:众所周知,所有现生棘皮动物的成体,均展现出规整的五辐射对称,而该门的部分幼虫形态却是两侧对称的。这条绝无仅有的个体发生(ontogenetic)特性,很可能暗示了一段充满波折的演化史:从两侧对称的祖先开始,终于辐射对称的冠群。长期以来,各种具过渡特征的化石纪录,一直被视作厘清其具体过程(辐射对称与其他衍征的整合顺序,以及每次转变的时间节点)的关键。尽管如此,“五辐射对称的起源”和“冠群最近共祖的形态”这两个论题,长期笼罩在不确定性的迷雾下:毕竟,纵观(已知)最早演化出步带的海旋板、较早显露出五辐射对称的海座星、始海百合(分别为游走类、有柄类的早期代表),此三者的形体构型似乎有天壤之别,使彼此间的同源性分析变得异常困难。然而,来自北非早寒武纪地层的Helicocystis moroccoensis,一种兼具海旋板(以Helicoplacus为例)与冠群特征的奇特化石,最终为这一难题的解决带来了曙光。
其一,Helicocystis的螺旋域、Helicoplacus的大部分体表③,均覆盖着螺旋排布的骨板。虽然两者的转动方向截然相反(前者顺时针、后者逆时针),却在间步带形成了完全一致、手凤琴样的页状伸缩结构。因此,Helicocystis的螺旋域,有可能同源于Helicoplacus的(整个)体壳。再结合前者的螺旋域与其余体段(萼+茎)间的分明界限,以及冠群棘皮动物体内的二分隔断(其两侧分别由被称为轴部、轴外部的幼体组织发育而来)。我们有充分的理由怀疑:Helicocystis的茎和萼,实际标志着“轴外部”构造的起源。
其二,Helicocystis的围口部,坐落在螺旋域的顶部,而其围肛部,则位于同一体段的侧面,而Helicoplacus的情况则恰恰相反。相比之下,同样营悬浮滤食(suspension-feeding)的有柄类(冠群棘皮动物),已经将口与肛门整合在萼的口面④。这或许意味着,消化道的扭转——一项发生于幼体固着后的变态过程中,并在棘皮动物的演化史上占据着重要地位的创新,很有可能经历了一个将肛门置于侧面的过渡阶段。
其三,Helicocystis的围口部,展现出接近于多种早期游走类(Stromatocystitidis、edrioasteroids)和有柄类(eocrinoids, glyptocystitids, diploporans,crinoids)的骨板布局,如果能证明这一性状的同源性,则不难得出:“七片骨板+“2-1-2”对称”的模式,很有可能是所有冠群棘皮动物的祖征。
其四,Helicocystis的固着部(茎/柄),虽然在形态上类似于某些原始的有柄类(如始海百合中的Gogia属),却会在个体发育中不断(相对其他部分)缩小。因此,有理由相信:在Helicocystis中,固着主要在幼虫末期的沉底-定居过程中起作用,对成体则不那么重要。从这种折中式的生存策略出发,有柄类的祖先保留并扩大了萼与柄,(幼态持续)、退化了其上的螺旋域,并将原本横卧体表的步带搬到了各种附肢上(arms/brachiole)。而游走类所在的世系,却选择让萼与柄进一步退化、成为位于螺旋域反口面中心的一小圈紧密镶嵌的骨板,彻底放弃了固着生活。
其五:在Helicocystis的步带底板上,并没有观察到通向身体内部的孔洞,因此,该属的水管系统可能为外置式(即卧于食物沟上方),且缺乏驱动管足的坛囊(ampullae)。然而,无论是比它更加古老的海旋板,还是从它分化而来的海座星,都具有类似的步带孔洞,暗示着棘皮动物在演化中多次获得-丢失这一性状的可能性。
简而言之,在已知的各种早期棘皮动物中,Helicocystis属展现出出人意料的性状组合(镶嵌):一方面是海旋板式的螺旋覆板,另一方面却是冠群棘皮动物的端位口、游走类的五条横卧式步带、以及有柄类的萼杯和茎/柄。因此,该属不仅填平了海旋板与冠群棘皮动物之间的深沟,更为冠群棘皮动物(游走类+有柄类)最近共祖的重建提供了重要参考。
注释(含个人理解)
①:原文指出,七片环口骨板中,仅有五片延伸到壳体的深处;如果剩下的两片位于C/D间步带,参考海座星的情况,有理由怀疑,它们可能由邻近的间步带骨板特化而来,以将原本被排斥在五片初始骨板之外的水孔/生殖孔整合到环口构造之内。
②:即五条步带中的四条(B与C,D与E)两两提前汇合,使直接通入口中的步带仅剩下三条,且呈“T”型布局。常见于五辐射对称的棘皮动物中的各种早期代表,如寒武纪的海座星和始海百合。
③:海旋板的骨板在近底部由螺旋排布转为竖直排列,似乎与固着有关。
④:原文中本无这一段(红字),仅为凸显“顶口侧肛”布局的过渡地位而补充。