
上一期,我介绍了泛霸王龙类的一些基础成员,它们通常是敏捷轻快的小型捕食者并在异龙超科的统治下艰难求生。土伦阶事件后,霸王龙超科迅速崛起,真霸王龙类统治的时代也随之而来。真霸王龙类(Eutyrannosauria)是泛霸王龙类下更加进步的演化支,定义是包含了君王霸王龙和鹰爪伤龙的最近祖先及其所有后代。
一、基础真霸王龙类恐龙

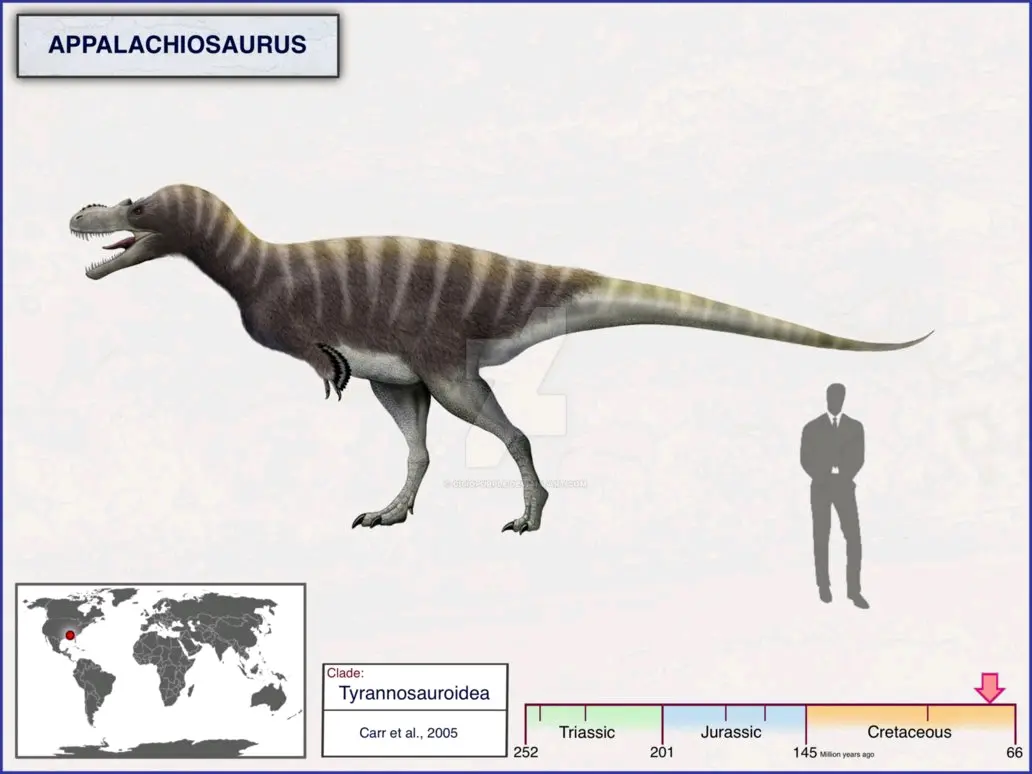
晚白垩世坎帕阶,阿巴拉契亚龙属,模式种蒙氏阿巴拉契亚龙,正模标本RMM6670在美国阿拉巴马州东部的白垩岩组被发现,包括头颅骨及下颌、几节脊椎、部分骨盆、及大部分后肢,均保存在阿拉巴马州伯明翰的麦克韦恩科学中心,该标本属于未成年个体,估测7米1.2吨。

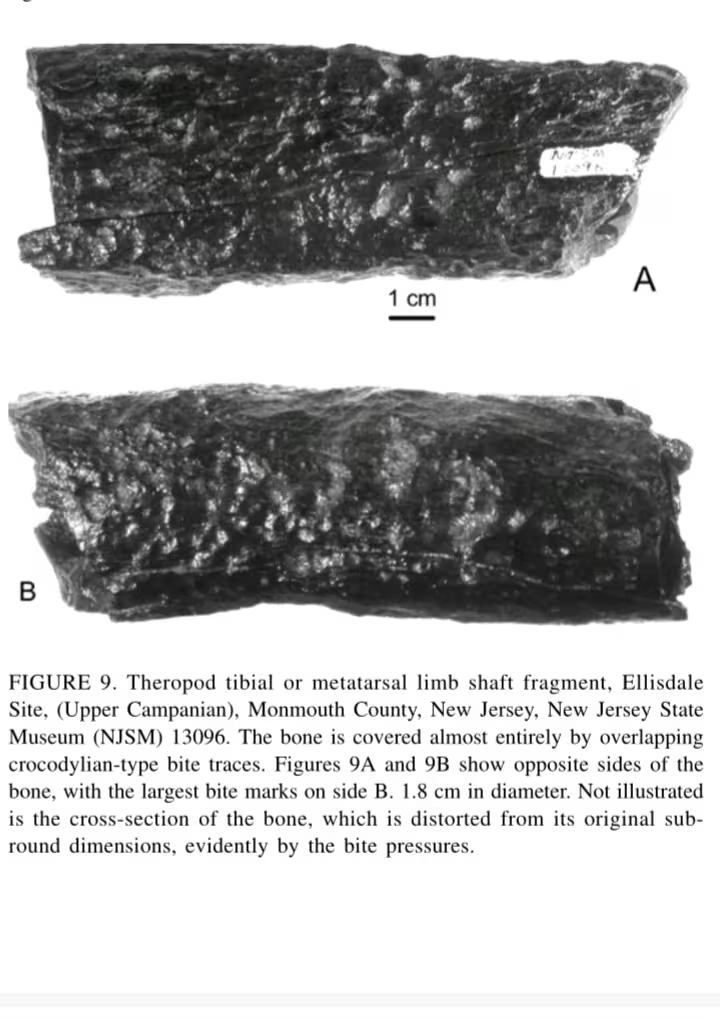
:一只300-400kg的兽脚类恐龙(可能是阿巴拉契亚龙幼年个体)的胫骨或者跖骨被褶皱恐鳄咬过,其骨头上留下最大的咬痕直径为1.8cm,这是目前唯一恐鳄攻击过掠食性兽脚类的证据。

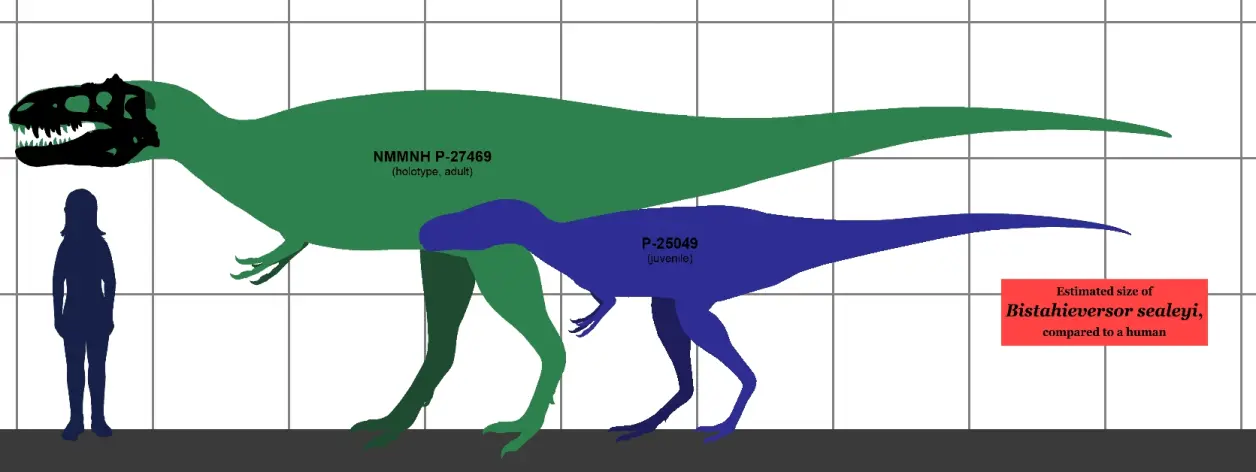
晚白垩世坎帕阶,虐龙属,模式种希氏虐龙,正模标本NMMNH P27469在美国新墨西哥州的科特兰(嘉德兰)组被发现,另有一具青少年个体NMMNH P25049,曾一度被归于阿尔伯塔龙属或者"后弯齿龙属"(疑名),正模标本估测8-9米2-3.3吨。虐龙属与五角龙属、结节头龙属共享栖息地。

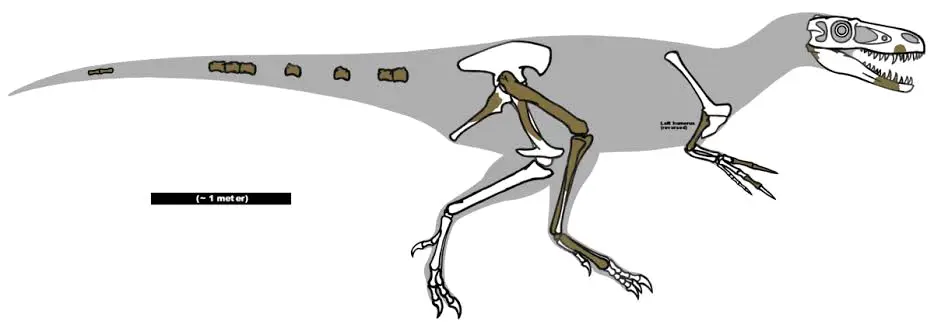
晚白垩世马斯特里赫特阶,伤龙属,模式种鹰爪伤龙,正模标本ASNP 9995发现于新泽西州的新埃及组,估测7.5米1.5吨,早期由爱德华·德林克·科普命名为"laelaps"(暴风龙),但这个名字已被一种厉螨占用,故被奥塞内尔·查理斯·马什于1877年更名为"dryptosaurus"(伤龙),种名"aquilunguis"意为具有老鹰的爪。伤龙曾被归入巨齿龙科(真是分类垃圾桶……),马什则为其建立伤龙科,后经与阿巴拉契亚龙比较,发现属于较为原始的霸王龙类。由于化石不完整,前肢趾爪的个数复原为2个或者3个。



二、霸王龙科恐龙
晚白垩世坎帕阶,血王龙属,模式种西南风血王龙,正模标本UMNH VP20200在美国犹他州南部的瓦维普组被发现,估测约为8米2.5吨,其属名含义是"血腥之王",种名则来源于古代希腊诗人荷马的西南风神,与已知最早的尖角龙亚科恶魔角龙属共享生境。西南风血王龙是目前已知最早的霸王龙科恐龙之一,其建属论文表明:霸王龙科是北美独立起源,而非先在亚洲早早诞生后迁入北美。血王龙的头骨侧面非常的高厚,颅后相当的宽阔,表明霸王龙类恐龙硬件演化的成功。

晚白垩世坎帕阶,力怖龙属(蛮霸龙),模式种统治者力怖龙,正模标本UMNH VP28348在2012年发现于新墨西哥州的梅尼非组,2018年正式叙述命名,包含部分颅骨、右侧第二掌骨、肋骨碎片、脊椎中部四个、左后肢第四根脚趾的两个指骨、骼骨,年代几乎和血王龙一样晚,估测体型为9米3.3吨,与结节龙科的无懈堡龙(Invictarx,模式种"西风无懈堡龙")共享栖息地。拉腊米迪亚的霸王龙类恐龙化石记录此前局限于7700万到6600万年前的K-Pg灭绝事件,而力怖龙的化石来自于比这早约300万年前的岩层,提供了来自坎帕阶早期的霸王龙类恐龙化石记录,将有助于古生物学家更好的了解霸王龙类恐龙的演化。

晚白垩世晚坎帕阶,怪猎龙属,模式种柯瑞氏怪猎龙,正模标本BYU8120在美国犹他州的凯佩罗维兹组被发现,并于2011年被汤马斯·卡尔(Thomas Carr)和汤玛斯·威廉森(Thomas Williamson)正式描述命名,属名含义为"怪物杀手",种名则来自于古生物学家菲利·柯尔(Philip J. Currie)。正模标本包含较完整头骨和大部分椎骨和肋骨、完整的骨盆、后肢部分,推测属于一只6米700kg的亚成年个体。
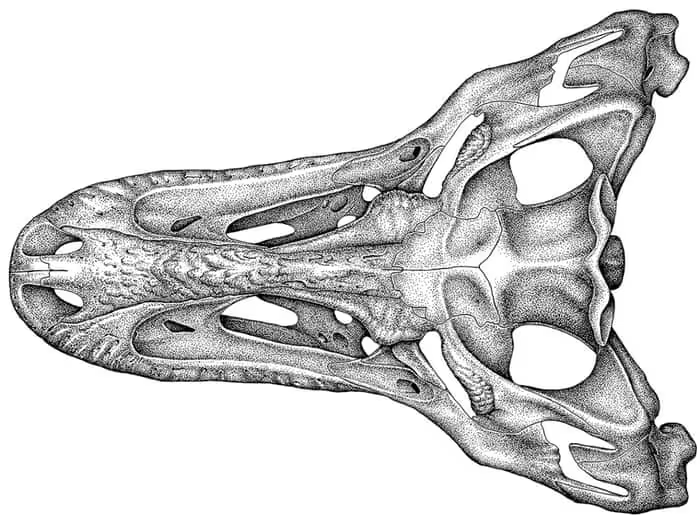
晚白垩世,马斯特里赫特阶,白熊龙属,模式种霍氏白熊龙,正模标本DMNH 21461在2006年发现于阿拉斯加的王子溪组,只有残缺的头骨,由美国古生物学家Anthony R. Fiorillo和Ronald S. Tykoski于2014年命名描述,根据高纬度的栖息地推测全长6米重达750kg,头骨的形状显示白熊龙的嗅觉非常发达。白熊龙与厚鼻龙属(陪罗特种)、"古植食龙属"和未定种伤齿龙科共享生境。

阿尔伯塔龙亚科:
晚白垩世坎帕阶,蛇发女怪龙属,模式种平衡蛇发女怪龙,正模标本NMC 2120于1913年在加拿大的恐龙公园组被发现,并于1914年被古生物学家劳伦斯·赖博(Lawrence Morris Lambe)叙述命名,估测能达到9.3米3吨,早期被怀疑和阿尔伯塔龙属是同一属甚至一度被归入阿尔伯塔龙属的一个种,但由于年代上的差异(蛇发女怪龙大致和阿尔伯塔龙相差4100万年),并且头骨仍有细微区别,所以蛇发女怪龙属目前仍然作为独立有效属。平衡蛇发女怪龙与强健惧龙共享栖息地,两种霸王龙科可能有着截然不同的食性和捕食技巧:前者可能更偏好手无缚鸡之力敏捷善跑的鸭嘴龙科且多在北部分布,后者可能更偏好有着较强防御、速度较慢的角龙科且多在南部活动。

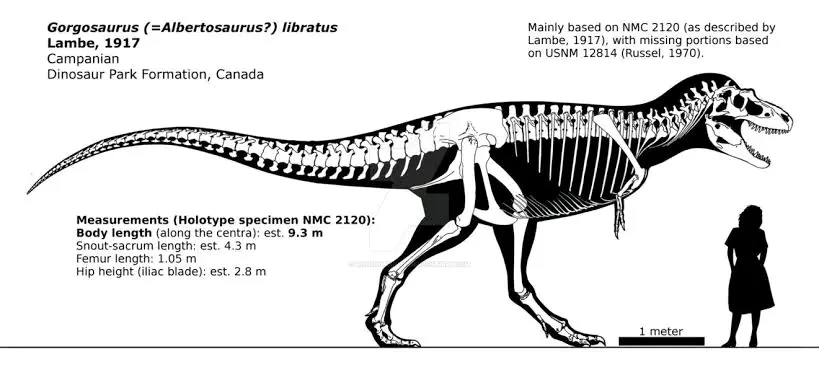
晚白垩世早马斯特里赫特阶,阿尔伯塔龙属,模式种肉食阿尔伯塔龙,正模标本CMN5600于1884年6月在加拿大阿尔伯塔省的马蹄峡谷组被发现,仅有部分头骨材料 ,估测全长9.3米3.2吨,在1905年被亨利·费尓费尔德·奥斯本(Henry Fairfield Osborn )叙述命名,属名是为纪念发现地加拿大阿尔伯塔省,意为"阿尔伯塔省的蜥蜴"。美国古生物学家巴纳姆·布朗(Barnum Brown)曾在干岛水牛躍省立公园处发现了至少9具肉食阿尔伯塔龙的标本,显示肉食阿尔伯塔龙拥有群居习性。
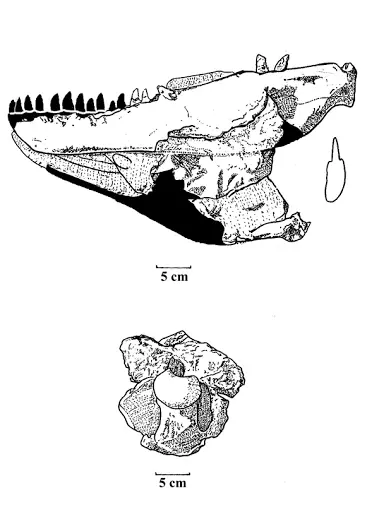

(?分类不定)晚白垩世晚马斯特里赫特阶,矮暴龙属,模式种兰斯矮暴龙,正模标本CMNH7541于1942年在美国兰斯组被查尔斯·怀特尼·吉尔摩尔(Charles W.Gilmore)发现,1946年被叙述并归入蛇发女怪龙属建立兰斯蛇发女怪龙种,1988年又被罗伯特·巴克 (Robert T. Bakker)及其同事重新叙述,认为该化石来自于成年个体(其实是),故建立新属—矮暴龙属,其含义是"侏儒暴君"。然而,后来的研究怀疑是霸王龙的幼年个体,2001年在蒙大拿州的地狱溪组又发现了一具名为"Jane"的个体(编号:BMRP 2002.4.1),可能是亚成年的矮暴龙,但由于标本问题也被归入了霸王龙。
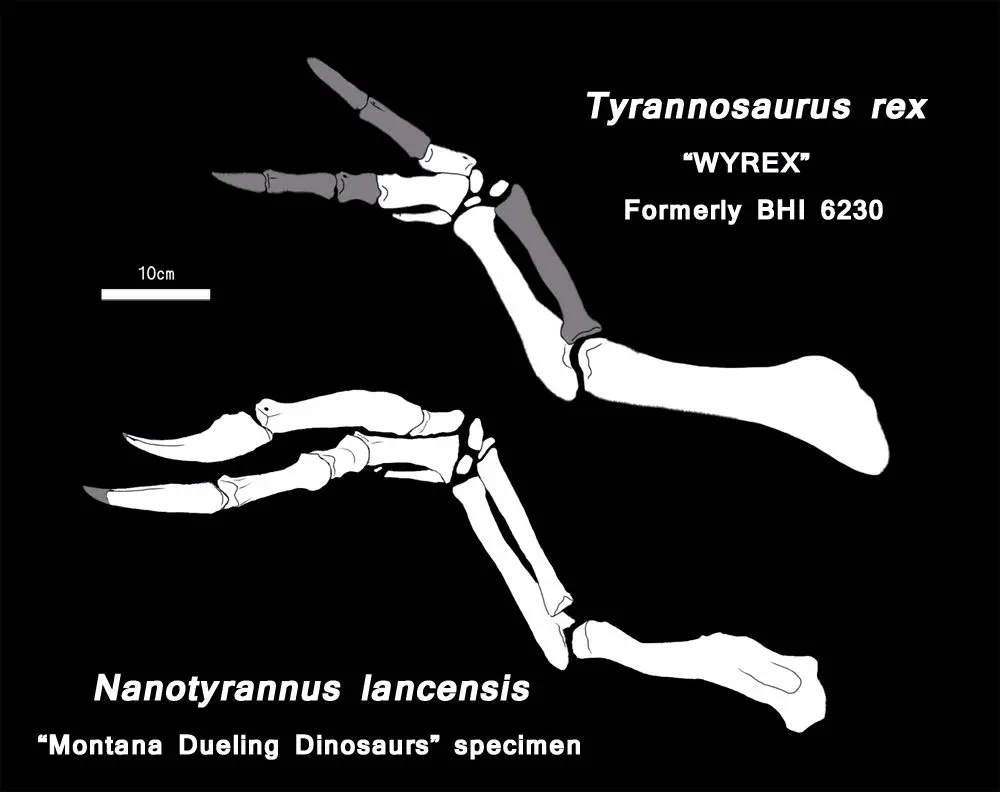
不过根据生长曲线,同等岁数的"矮暴龙"和霸王龙体型相差非常大,举个例子:矮暴龙的正模标本年龄估测为8岁但却只有5.3米520kg,而与之同岁的霸王龙理论可能有7-8米2吨;年龄估测为11岁的"Jane"估测6.5米960kg,而此时的霸王龙理论已经有9-10米4吨的大小,显然两具标本是非常不符合霸王龙生长模式的【详情见恐龙吧洪穹神皇上帝2018年的帖子霸王龙生长模式(新版:~99%无矮版)】。另外,矮暴龙上颌骨单侧有17颗牙齿,而成年霸王龙上颌骨单侧牙齿数为14颗,矮暴龙的前肢也远大于霸王龙,见下图:
综上所述,矮暴龙属理应是有效的,并不是某些学者声称的所谓矮暴龙标本全是未成年霸王龙,但奈何霸王龙缺乏幼年个体的标本做更进一步的对比研究,因此矮暴龙的头骨独征也被反对派认为是霸王龙幼年个体和成年个体的发育差异,至今学术界仍有一部分争议。
此外可能的,矮暴龙标本还有私人收藏标本"血腥玛丽"(幼年),BHI662"Chomper"5(幼年),"Petey"(成年,暂未查到编号,活体估测7米1.5吨)。另外,曾被认为是青少年个体或成年矮暴龙的LACM23845,很可能是地狱溪组第三种霸王龙类"恐暴龙(Dinotyrannus)"。
矮暴龙和"恐暴龙",伤龙,以及新泽西州纳维锡克组出土的未定种基础真暴龙类恐龙(编号MAPS A 1210a/b,化石的主人可能有9-10米)的存在表明晚白垩世马斯特里赫特阶的北美洲恐龙仍然拥有较高多样性,而不是完全走向衰败。

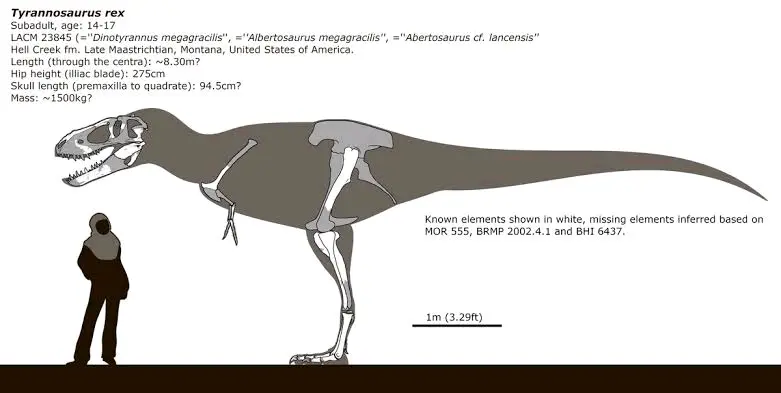
本文最后一部分,霸王龙亚科:
分支龙族:①晚白垩世,虔州龙属,模式种中华虔州龙,正模标本在中国江西省赣州市的南雄组被发现,头骨全长90cm,估测大小为6.5米800kg,化石包含完整的头骨、大部分颈椎、较完整后肢、部分背椎和尾椎,于2014年5月公布于《自然杂志》。中华虔州龙狭长又极窄的吻部表明它与蒙古的分支龙属有着很近的亲缘关系,因此建立分支龙族演化支,也得了"匹诺曹暴龙"的称号。虔州龙属与赣州龙属、南雄龙属、通天龙属等共享生境。
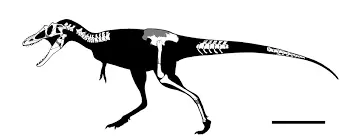
2019年7月,江西发现的霸王龙类恐龙脚印的主人很有可能是虔州龙属,脚印长度达58cm宽47cm,造迹者估测7.5-9米,远比虔州龙正模大。

②晚白垩世马斯特里赫特阶,分支龙属,模式种遥远分支龙,正模标本PIN 3141/1在蒙古巴彦洪卓省的尼古特萨(Noggon Tsav)组被发现,包含大部分头骨、三个相连跖骨,于1976年被俄罗斯古生物学家谢尔盖·库扎诺夫(Sergei Kurzanov)叙述命名,该标本头骨低矮狭长、矢状脊发达、上颌牙齿数(16颗)比其它霸王龙科恐龙多,与很多霸王龙科恐龙不同,因此被命名为"遥远分支龙",估测5-6米,不过头骨化石在形成过程中变形,可能比原先预计的更短。另外,正模标本应该是未成年个体,成年个体会更大。

分支龙属还有一个种,阿尔泰分支龙,正模标本IGM 100/1844在蒙古纳摩盖吐组被发现,化石比模式种更为完整,由斯蒂芬·布鲁萨特(Stephen L. Brusatte)等人于2009年叙述命名,估测,估测5米450kg,死亡年龄9岁,未成年个体。
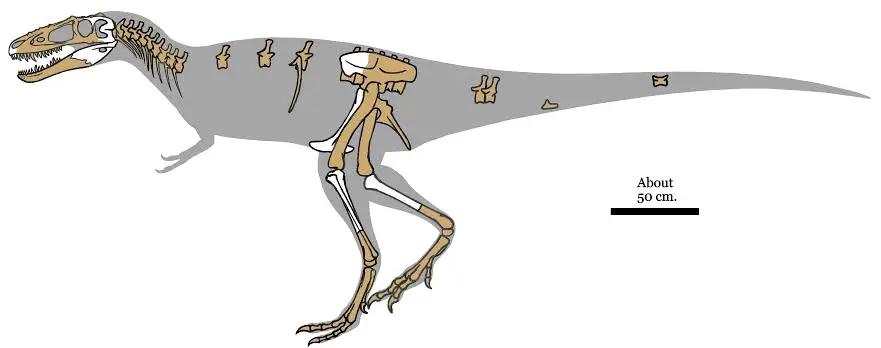

惧龙族:①晚白垩世晚坎帕至早马斯特里赫特阶,惧龙属,模式种强健惧龙,正模标本CMN8506于1921年在老人组被查尔斯·斯滕伯格(Charles Mortram Sternberg)所发现,估测9.8米4.5吨(活体预计),早期被认为是蛇发女怪龙属的一个种,直到1970才被戴尔·罗素(Dale Russell)完全叙述,建立“惧龙属”,属名“daspletosaurus”其含义是“令人畏惧的蜥蜴”,种名“torosus”则在拉丁文中指“强壮”或“结实”,该标本估测。

惧龙属的另一个种霍氏惧龙(晚坎帕阶),正模标本MOR590在美国蒙大拿州的双麦迪逊组被发,包含完整的头骨、部分前肢和近乎完整的后肢,估测9米3.4吨,于2017正式描述,种名以美国著名古生物学家兼前蒙大拿州博兹曼市的洛基山博物馆馆长杰克·霍纳(John'Jack'R. Horner)命名。与它的姐妹物种强健惧龙不同的是:强健惧龙的泪腺支比腹枝更长,表明霍氏惧龙的头骨更高,强健惧龙的泪眶窝被腹枝的一个深凹隔开,而这些表面在霍氏惧龙上是融合的,强健惧龙外侧面和背侧面的冠状区域相等而霍氏惧龙则是外侧面多于内侧面。
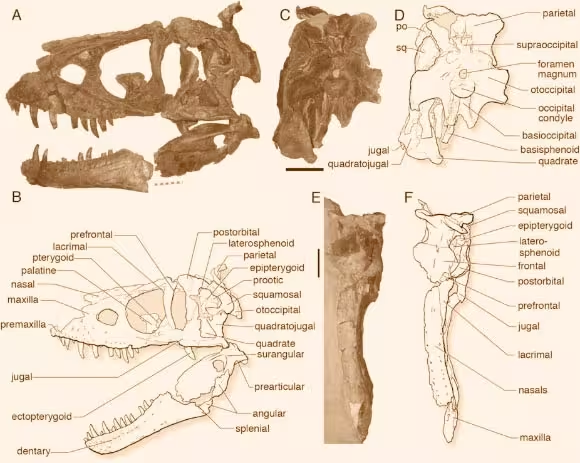

除此之外,还有惧龙属未定种:
恐龙公园组—代表标本为1914年被发掘的标本FMNH PR308,曾长期置于"平衡阿尔伯塔龙"中;
嘉德兰(科特兰)组—代表标本为OMNH 10131,,被怀疑是虐龙属的材料;
朱迪斯河组—RMDRC 06-005,惧龙属的最大个体,化石材料只有残缺头骨和零碎身体骨骼,头骨全长预计136-140cm,活体全长预计10.8米体重6吨,年代估测为晚白垩世晚坎帕阶,该标本挖掘清理用时近10年。
马蹄峡谷组—CMN 113513,亚成年个体,估测年代是晚白垩世早马斯特里赫特阶。
②晚白垩世,死掠龙属,模式种德氏死掠龙,正模标本TMP 2010.5.7在加拿大阿尔伯塔省的Foremost组被发现,化石仅包含部分头骨碎片,估测8米2吨,不过该标本未停止发育,是亚成年个体。2020年2月被正式描述命名,属名来源于希腊神话中的"死神",含义是"死亡收割者",而种名则是对发现者加拿大艾伯塔省的约翰·德·格鲁特(John·Sandra ·De·Groot)的致敬。德氏死掠龙发现意义重大,它填补了我们对霸王龙进化理解的空白,带来一个全新的演化支—"惧龙族"。德氏死掠龙与角龙类的异形角龙属和肿头龙类的结节头龙属共享生境。
霸王龙族:
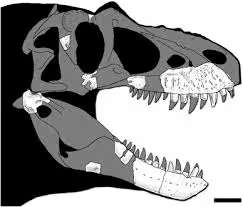
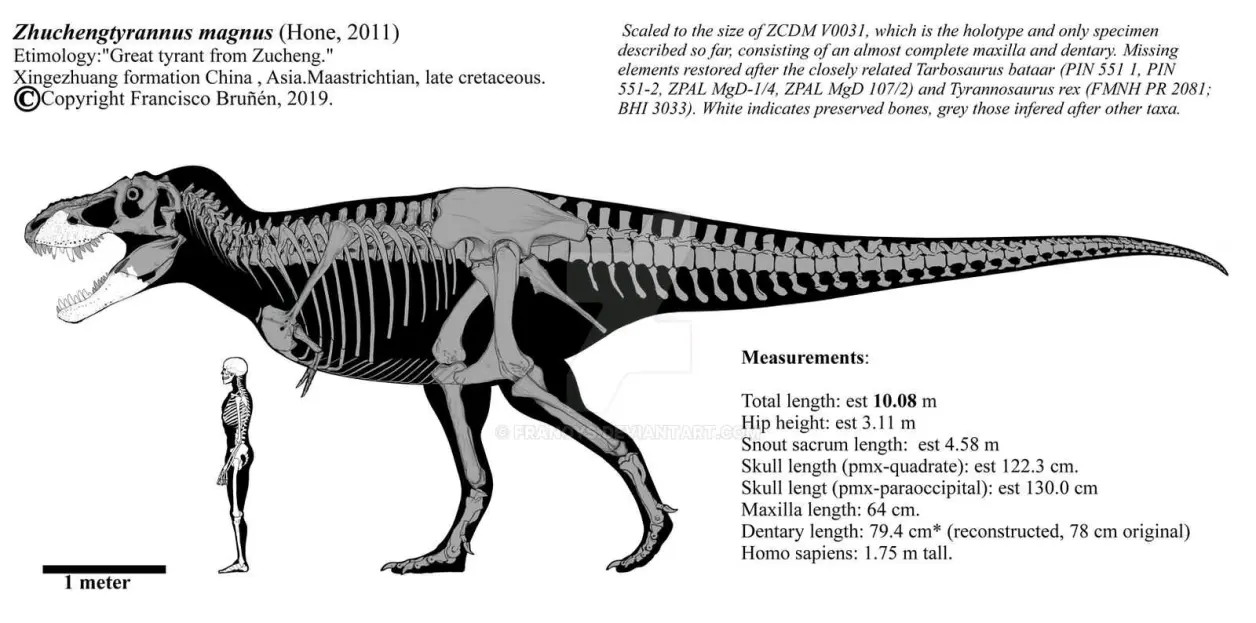
①晚白垩世晚坎帕阶,诸城暴龙属,模式种巨大诸城暴龙,正模标本ZCDM V0031在中国山东省辛格庄组王氏群被发现,包含完整的右上颌骨和左齿骨,估测10.6米5.8吨(活体预计)。早期被置于霸王龙属里("诸城霸王龙"种),因为诸城暴龙头骨形态是最接近霸王龙的,不过诸城暴龙上颌骨上升突基部有突起,上颌窗孔前缘有一圆形凹痕,及一些独特的特征组合,区别于霸王龙类的其他所有属种,因此于2011年建立"诸城暴龙属(Zhuchengtyrannus)",这一发现为中国的古生物学带来了不小的热度。
诸城暴龙属有个疑名"瘦长诸城暴龙",于2011年5月被中国科学院古脊椎动物与古人类研究所著名恐龙专家徐星发现,缺乏描述,暂无详细信息。
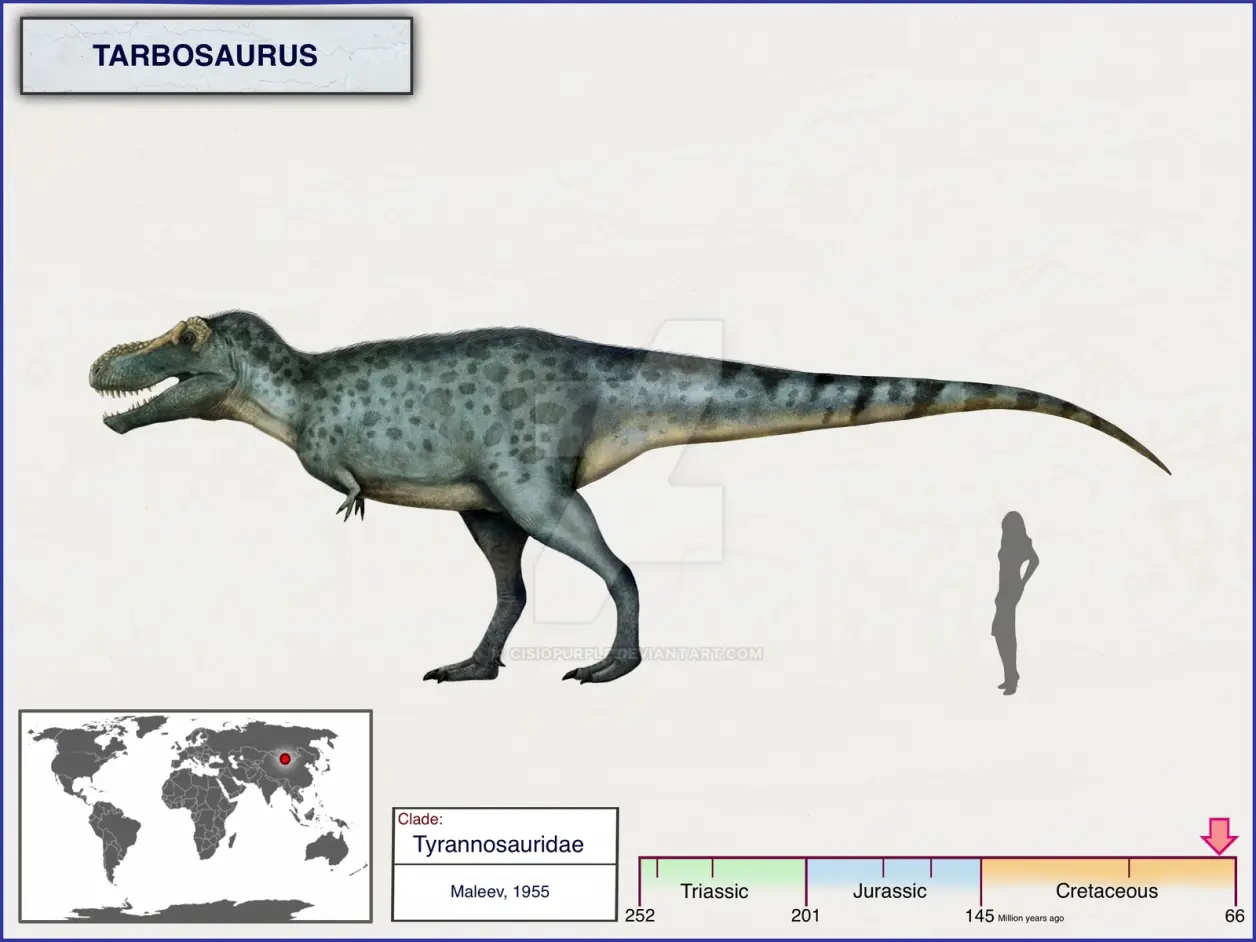
②晚白垩世,早马斯特里赫特阶,特暴龙属,模式种勇士特暴龙,正模标本PIN—555—1于1946年被一个苏联与蒙古挖掘团队在蒙古纳摩盖吐组发现,包含较完整头骨和部分椎骨,估测11.2米6.5吨(活体预计),1955被苏联古生物学家叶甫根尼·马列夫划分在霸王龙属里("勇士霸王龙"),同时将其它被发现的标本归于蛇发女怪龙属的不同种。之后1956年,苏联古生物学家阿纳托利·康斯坦丁诺维奇·罗特杰斯特文斯基认为马列夫的标本是同一种动物不同生长阶段,并不同于霸王龙属,故新建"特暴龙属(Tarbosaurus)",之后不少专家也认同该分类。
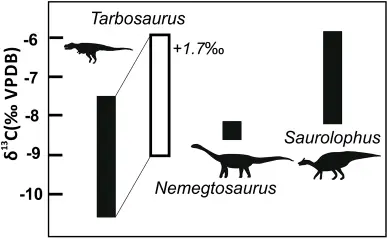
2003年,一项有关特暴龙头骨的研究显示:当特暴龙咬住猎物时,力量直接从上颌传递到上颌周围的颅骨,这一点与北美的霸王龙科恐龙不同,北美的霸王龙科咬合时力量传递到口鼻部上方的固定鼻骨再经鼻骨与泪骨间的骨质连接至泪骨。特暴龙的鼻骨缺乏与肋骨间的骨质连接,并且隅骨侧棱脊连接齿骨后方的方形突,这使得特暴龙的下颌无法灵活内外扳动,这与其它霸王龙科恐龙区别,因此特暴龙的这种坚硬结构很可能是为猎杀纳摩盖吐组的蜥脚类(如:长达12米的纳摩盖吐龙)而演化。
2019年,一项关于特暴龙饮食的研究一定程度证实了该理论:研究人员提取了出土于纳摩盖吐组的5个不同的特暴龙个体牙齿的样本,并与同地区的其他素食恐龙(包括纳摩盖吐龙、栉龙、巴思钵体龙等)的牙齿和骨骼样本进行了对比,结合已有化石咬痕,发现特暴龙更倾向于捕食鸭嘴龙科的栉龙和蜥脚类,同地区的杂食性大型兽脚类也是潜在的猎杀目标。
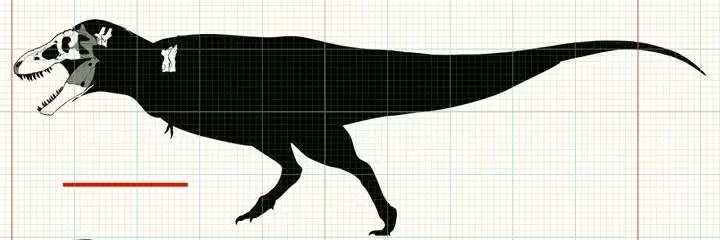
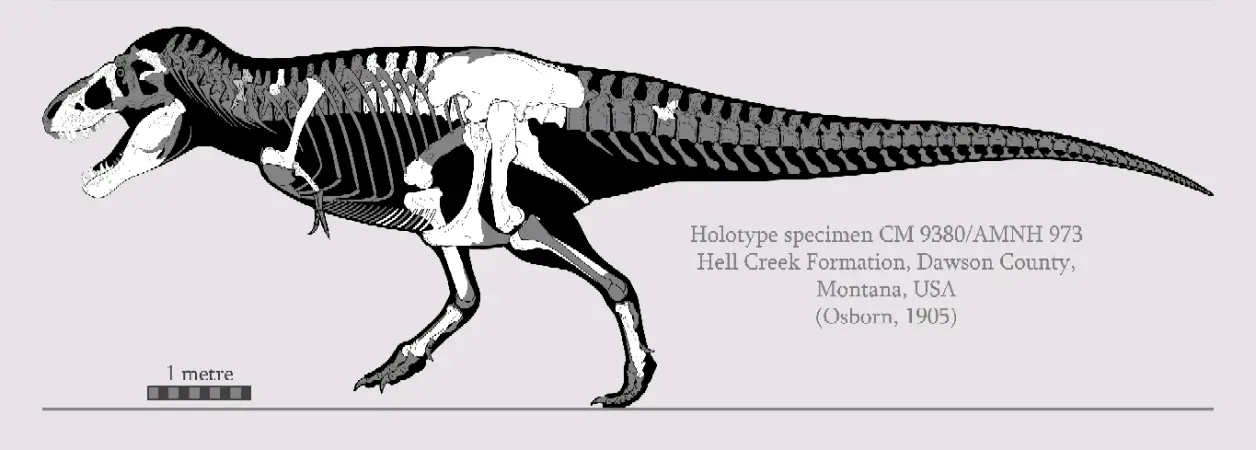
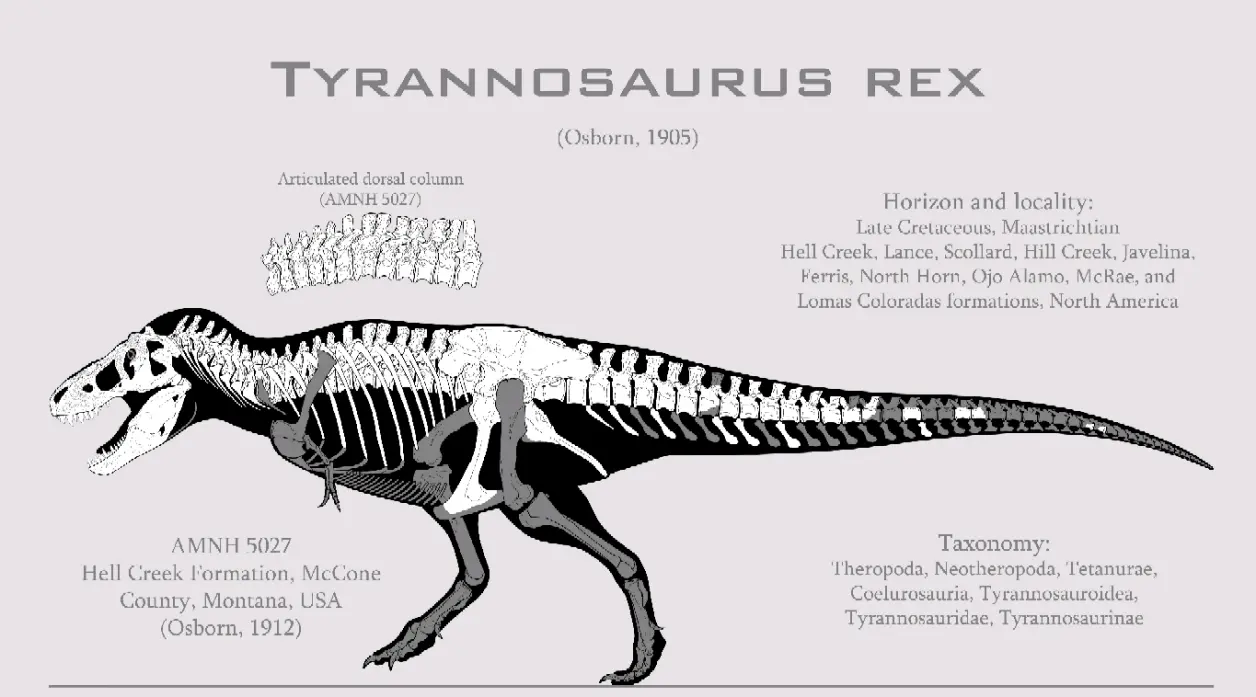
③晚白垩世晚马斯特里赫特阶,霸王龙属,模式种君王霸王龙,正模标本CM9380(曾用编号:AMNH973)于1902年被巴纳姆·布朗(Barnum Brown)在美国蒙大拿州的地狱溪组被发现,该标本体型估测(活体)12.4米9.7吨,布朗随即把这个消息告诉亨利·费尔费尔德·奥斯本(Henry Fairfield Osborn )。3年后奥斯本将其正式描述命名为"君王霸王龙(Tyrannosaurus rex)"时,由于当时人们对掠食性恐龙的认知局限于侏罗纪的兽脚类,所以正模的短前肢便遭到怀疑可能是将一只较小的与另一只较大的恐龙化石混杂,并且正模标本那残缺的头骨用异特龙的材料进行修补。随着1941年美国加入二战,正模标本被卖给了匹茨堡的卡内基自然历史博物馆,编号改为CM9380。最初,它的装架设为袋鼠站式,后经现代研究被修正并且给它更科学的头骨重建。
说到CM9380的头骨重建,就不得不提一下AMNH 5027:它同样是由巴纳姆·布朗与1908年在美国蒙大拿州的地狱溪组呗发现,于1912年到1916年间被奥斯本描述,推测体型与正模相近。正是这个标本的发现,使得霸王龙短粗、结实的脖子被曝光,还提供了完整的头骨,使得CM9380的头骨得以修正。AMNH5027与之后发现的标本(如BMNH R7794和FMNH PR2081)相比更,因此随着后来的发现霸王龙与其它肉食龙的脖子区别越来越明显。AMNH5027的意义不仅仅局限于科研,它还是大名鼎鼎的科幻冒险电影《侏罗纪公园》系列里霸王龙的原型,在影视史上留下了浓墨重彩的一笔。
虽然AMNH5027不是最大的霸王龙,但它绝对是影响最大意义最深的那只霸王龙。
AMNH5027还是UCMP118742最适合估测的模板,因为UCMP118742的上颌骨残片最接近AMNH5027,那么按照AMNH5027放大,活体预计将会达到惊人的14.3米14.8吨。

霸王龙真正的第一具标本是1892年被爱德华·德林克·科普所发现的AMNH3982,只有两个脊椎骨(其中一个已遗失),最初被他认为属于角龙类,被命名为"大薄椎龙(Manospondylus gigas)",意思是"巨大的多孔椎骨",该标本后来被鉴定为是属于兽脚类而非角龙类恐龙的标本,而早在1917年,亨利·费尔费尔德·奥斯本(Henry Fairfield Osborn ) 就意识到"大薄椎龙"与霸王龙骨骼的相似性,只是由于椎骨的碎片性奥斯本未将该标本归入霸王龙属。该标本疑似是14米级别的超大个体。
不过至今为止,学术界公认最大的霸王龙是FMNH PR2081"苏",已知最完整的霸王龙标本,学术界主流采取的体型估测是12.35米8.8吨(活体修正12.8米11.7吨),于1990年月日在南达科他州的地狱溪组被发现,随后卷入了关于其所有权的法律战争(这里不详细描述了,感兴趣的朋友可以去外网查询)。据推测,苏巨大体型可能是因为长期的个体发育,因为它是已知年龄第三老的霸王龙,据彼得·麦克维奇和佛罗里达大学的估计,苏死亡年龄为28岁,比大多数霸王龙标本(如MOR555、AMNH5027、BHI3033)还大6-10岁以上,已知的所有霸王龙标本只有Trix和Scotty比苏更老。苏凭借着73%+的完整度一举斩获最大霸王龙以及最大兽脚类的名号,据称比苏大的霸王龙,往往由于自身完整度和研究不深入的问题并未被承认。

所以不要迷恋苏,苏不只是传说~~~(滑稽)
关于霸王龙属是否具有羽毛还有待进一步的研究,霸王龙属有皮肤化石发现白垩纪气候热霸王龙体型大≠完全没有羽毛,只是会非常稀疏。还有已发现羽毛的霸王龙超科只有帝龙属和羽王属龙,这俩都是原角鼻龙科的,我之前的专栏提到泛霸王龙泪是除了原角鼻龙科(可能还有大盗龙类)以外的霸王龙超科,它们的关系离霸王龙都很远,并且早白垩世的亚洲地区是偏寒冷的,这基本不能作为霸王龙属批羽的证据。

而且霸王龙不是骨骼密度最高的恐龙,骨骼密度极高往往是特化水生动物的特征,之前动态的对比图再发一次

另外关于霸王龙是否食性问题:
从下图可以看出,地狱溪组数量最多的动物是三角龙属,地狱溪组是霸王龙属主要分布的地层,在这里鸭嘴龙科的数量比三角龙属显得少得多,这完全可以说明君王霸王龙会以该地区的褶皱/前凸角三角龙为主食 而不是完全选择毫无还击手段的鸭嘴龙类。
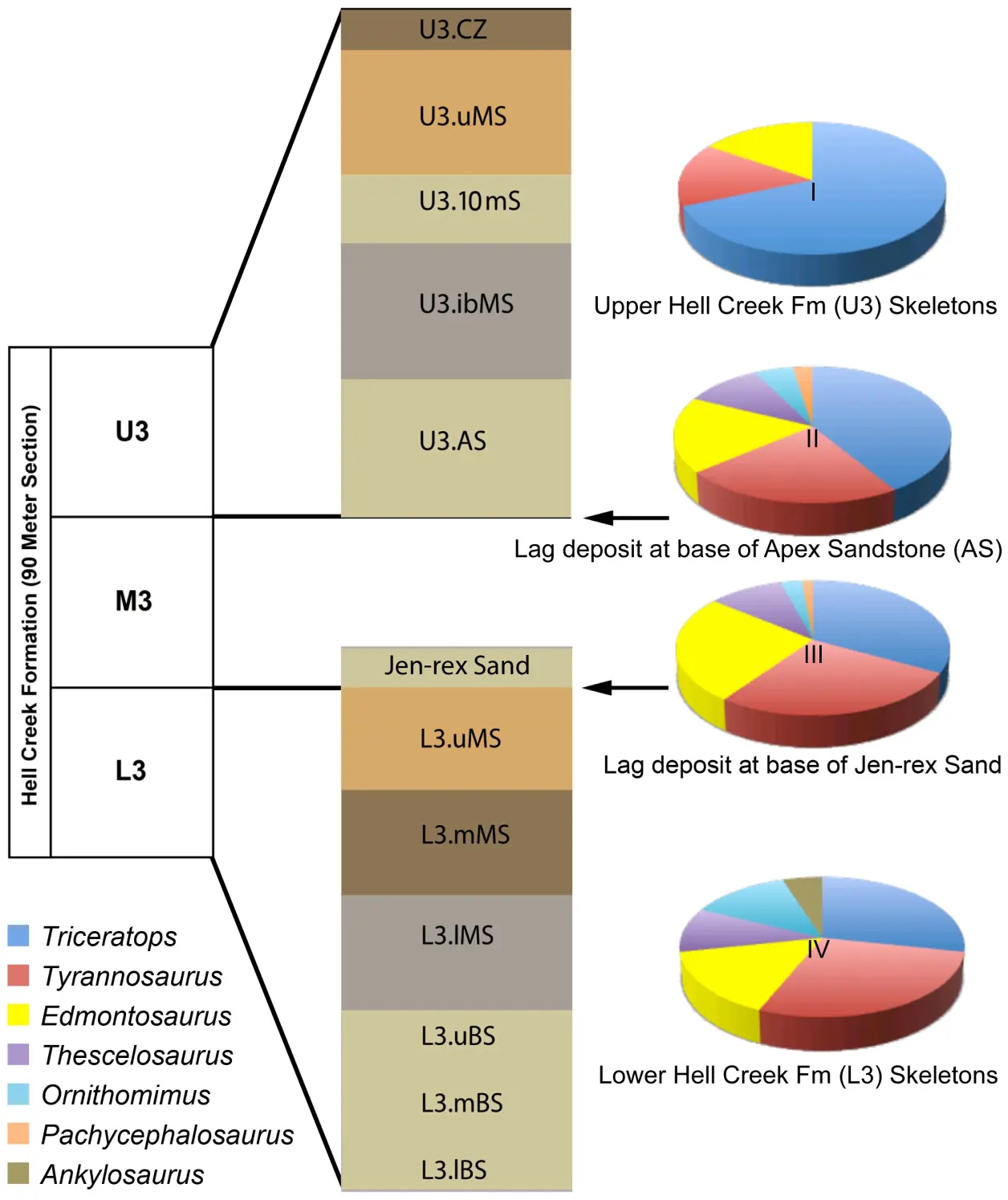
饼图I-IV反映了基于骨骼(图I和图IV)和个体骨骼(图II和图III)的恐龙属的相对丰度。霸王龙的骨骼在上地狱溪地层中的数量与食草动物鸭嘴龙科的埃德蒙顿龙一样丰富,在下地狱溪地层中的数量几乎是埃德蒙顿龙的两倍。霸王龙和埃德蒙顿龙的个体骨骼在滞后沉积物中发现的数量大致相同。注意:饼图一,上地狱溪组(U3)中的恐龙骨骼;饼图二,顶点砂岩(AS)底部“Doldrum's”滞后沉积物中的恐龙骨骼;饼图三,Jen rex砂岩底部“3B-1”滞后沉积物中的恐龙骨骼;饼图四,下地狱溪组(L3)的恐龙骨骼。缩写:L3lBS,下部基底砂岩;L3.mBS,中部基底砂岩;L3.uBS,上部基底砂岩;L3.lMS,下部泥岩;L3.mMS,中部泥岩;L3.uMS,上部泥岩;U3.AS,顶部砂岩;U3.ibMS,互层泥岩;U3.10mS,10米砂岩;U3.uMS,上部泥岩;U3.CZ,煤田。
生态平均霸王龙与三角龙的体型比例大致是11.5米7.6吨:7米5.5吨,极端上限的体型比例则是14.3米14.8吨(编号UCMP118742):8.2米8.6吨/9.2米10吨(编号USNM8018/UCMP128561,霸王龙是完全占据体型优势,不需要田忌赛马。
可能会有人说:"三角龙的尖角完全能刺死霸王龙呢!而且三角龙比霸王龙灵活!!!"
转神皇:计算霸王龙速度的研究和对应的子模块(作为计算应用方法所需要的参数模块),霸王龙数据远远比三角龙全面,因为霸王龙研究全面了,三角龙很多数据停留在70~80年代,有时候会比较头疼,这里霸王龙64~104km/hr,三角龙45~55km/hr这些一看就是卫星的就不用拿出来...当然比卫星也是霸王龙赢。用几种霸王龙和三角龙都适用的方法
【方法1】假设两者都不能肢体同时间全部离地,那么按两者最快的步姿,骨骼最大承受力量是多少,霸王龙步姿很简单,如果不能双足离地,那就是一只脚落地,三角龙最快的步姿是非对称跑步,一足着地,三足离地。这种情况下谁骨骼承受力强,谁单肢产生的反作用力就更大,速度就更快。三角龙的股骨是最薄弱环节,霸王龙的股骨和跖骨并列最薄弱环节,因此大家都测算股骨
先说双方大个体(大家都喜欢大个体吗)。
1)霸王龙Scotty体重11~12吨,股骨周长590mm,股骨长度Lc-H 1370mm(Lc-Gt 1333),Lm是199mm,AP(修复后,按椭圆指数倒算)是175mm,股骨前后运动强度指数(Lm*AP^2)/L=(199*175^2)/1370=4448,综合体重后指数为370~404
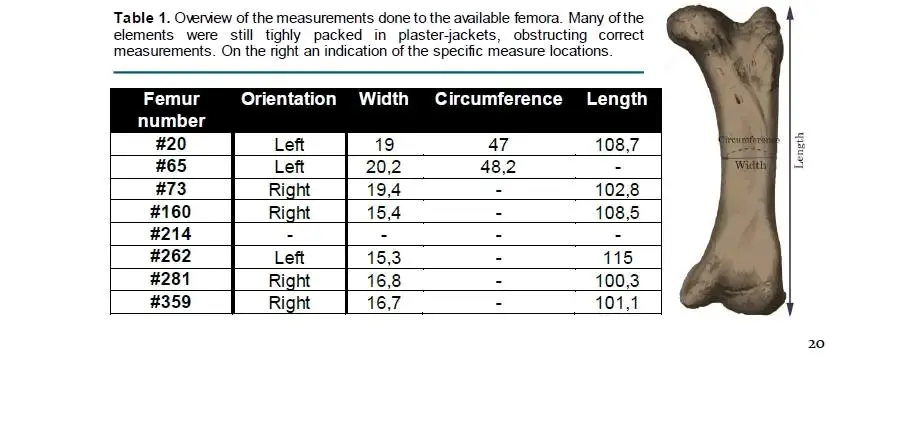
2)三角龙体重7.2~8.4吨(非完整骨架,缩放不同个体的差距),LACM 150076股骨长度1171mm,周长533mm,Lm按下图的#20三角龙,Lm是215.4mm,AP是117mm(按椭圆指数倒算)~124.5mm(参考#65的切面),股骨前后运动强度指数=(215.4*117*124.5)/1171=2679,综合体重后的指数为319~372
3)霸王龙再选一个股骨比较细的一般个体,BHI 3033(按小STAN版),体重8.6吨,股骨LM 168mm,AP 139~150mm(前者实测但是略破碎断面,后者是椭圆指数;MOR 555/8.2吨,股骨1280mm,LM 165mm, AP 146mm可以作为参考,虽然也碎),股骨强度指数2740,综合体重后为319
4)霸王龙再选一个股骨比较粗的一般个体,CM 9380,体重9~10吨,实测股骨LM 182.5mm, AP 161.5mm,股骨长度1278mm,强度指数是3724,综合体重后是372~414
5)三角龙#20,体重按5.8~6.7吨,周长是470mm,股骨长度1087mm(小结高度,但和最大长相等) LM和AP是190mm、103mm,强度指数是1854,综合体重后是277~320
6)三角龙#65,体重按6.2~7.2吨,周长是202mm和117mm,或者按96mm(按椭圆指数),股骨长度1114mm(参考#20),强度指数是2036,综合体重后按283~328
7)霸王龙再选一个小个体(11岁,刚成年或者准成年),MOR 009,这个是6吨,和平均成年三角龙(5.3吨)很接近,股骨长度是1150mm,周长469mm,股骨AP 112mm(实测),LM 150mm(实测),指数1636,综合体重后是273。
8)亚成年个体USNM 6183,4吨左右,股骨周长425mm,股骨长度Lc-Lt 990mm(估测Lc-H1040mm),LM 149mm, AP 120mm,强度指数2063,综合体重后515。
10)三角龙里面也选个比较细的,例如1150mm的,大概6,4吨(=USNM 4812)LM 153, AP参考#20的比例也就88mm,强度指数(153*88*88)/1150=1030, 综合体重后指数是161
11)再按下图最小的#281的股骨,股骨长度1003mm,体重大概是4.25~5.3吨,LM=168, AP96.6,强度指数1563,综合体重后是295~367
12)按下图相对最粗壮的三角龙股骨#73,这个个体是股骨长度1028mm,但是Lm 194mm,估测AP 111.5,估测体重是4.9~5.7吨,考虑股骨很粗,此处选择按5.7吨,指数2346,指数在411左右。
结论就是总的来说霸王龙前进的速度至少不小于三角龙,霸王龙1), 3), 4), 7), 8)取平均是387,,319,393,273,515;三角龙是345.5,305.5,298.5,161,331,410,;霸王龙对三角龙占优势(一些老论文说三角龙Strength Indictor比霸王龙高,此处数据结论不支持),当然三角龙(161~410)和霸王龙(273~515)有一定的重叠。
这里说的是前后前进强度,如果是撑地转弯强度,公式是LM^2*AP/L,则以上的霸王龙1), 3), 4), 7), 8)分别是440,371,444,366,639;三角龙则是616,484,550,280,575,713,总的来说转弯上三角龙占优势。但是同样有一定的重叠。
这里只转一部分,结果已经显而易见了:三角龙对霸王龙没有绝对优势,至于反杀吗?看非洲草原被水牛整死的狮子多少咯???现今其它掠食者被素食动物反杀的概率多高呢???这种问题,不需要梗深层次讨论了,没有绝对以腐肉为食的脊椎动物,当然霸王龙摔死以及被同类杀死的概率是比三角龙反杀更高的。
不要以斗兽一定争强弱的思想看自然生态,毕竟不是游戏平衡玩家数值……霸王龙是强大的掠食者没错,但也是曾生活在地球上千万生命的一份子,与其它生物没有绝对的贵贱之说。望诸君客观看待,切忌无脑斗兽。
泛暴龙类演化支的故事到此结束,感谢阅读。
总结:霸王龙超科是相当成功的类群,从中侏罗世巴通阶的布氏原角鼻龙到晚白垩世马斯特里赫特阶的君王霸王龙,它们或大或小并拥有各自的生态位,历经100多万年诞生了史上最大的陆地掠食者,虽然仅是昙花一现,但不可否认在地球史上留下了生命的光辉,一定程度向我们展示了恐龙总目的多样性,正如同其它兽脚类恐龙一样。
如有错误,欢迎指正!
参考资料:
维基百科
https://phys.org/news/2019-06-tooth-enamel-analyses-insights-diet.amp(特暴龙饮食研究)
https://www.nature.com/articles/srep44942(霍氏惧龙论文)
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0079420(血王龙相关论文)
https://tieba.baidu.com/mo/q/m?kz=5824175572&from_search=1&sc=121246132022&lp=se_thread_pick_&pn=0&(霸王龙生长模式,帖子较老,矮暴龙部分请排除LACM23845和某AMNH个体)
https://tieba.baidu.com/p/5660402447(霸王龙感官)
https://tieba.baidu.com/f?kz=5677487666&mo_device=1&ssid=0&from=1022282j&uid=0&pu=usm@1,sz@320_1001,ta@iphone_2_8.1_24_62.0&bd_page_type=1&baiduid=1D3AF09D532F04C891E5158977A6147E&tj=www_normal_1_0_10_title&referer=m.baidu.com(霸王龙速度探讨)