
用于分析神经活动的新一代光遗传学工具有助于阐明神经科学中的经典开放问题。具体而言,电压成像技术,使用增强的基因编码的电压指标已越来越多地用于观察大电路的动态中尺度。在这里,我们描述了如何结合联合收割机皮层宽场电压成像与海马电生理清醒,行为小鼠。此外,我们强调这种方法可以是有用的不同的可能的调查,使用的表征作为一个案例研究的大脑皮层的相互作用。
图片上传处理中...
东莞富临医疗科技有限公司是Open Ephys 和 Intan Technologies 在亚洲的代理商,富临医疗为亚洲客户提供“技术服
务”与“电生理产品”
一、引言
海马体-皮层相互作用已被广泛研究,旨在理解其在支持记忆中的作用。然而,大多数研究集中在海马体与特定皮层区域之间的相互作用上,这些区域与海马体有主要的解剖连接,例如内嗅皮层或内侧前额叶皮层。关于海马体活动与跨越大部分皮层空间的活动现象之间的相互关系知之甚少。皮层范围的活动通常通过依赖神经活动间接相关信号的成像技术进行研究,例如血氧水平依赖(BOLD)信号或细胞内钙信号,而我们对海马体的了解主要来自微电极记录。虽然经典成像方法具有出色的空间覆盖范围,但其空间分辨率有限。另一方面,微电极技术提供了具有出色时间分辨率的记录,但空间信息有限。特别是,使用基因编码钙指示剂(GECIs)的传统宽场成像在活动时间上与电生理学存在显著差异,这是由于记录信号的性质不同。在头部固定的小鼠中使用基因编码电压指示剂(GEVIs)进行皮层宽场电压成像,可以提供具有高空间和高时间分辨率的皮层活动大尺度视图,并且可以与基于电极的电生理学结合使用。本文描述了将皮层电压成像与海马体电极记录相结合,以同时监测休息和传统行为任务期间的皮层和海马体活动。具体而言,我们举例说明了这种组合如何有助于解决感觉-海马体整合和海马体-皮层相互作用相关的问题。
二、结果
2.1 同时进行皮层电压成像与海马体电生理记录
为了研究海马体与皮层之间的相互作用,我们使用了皮层宽场电压成像与海马体CA1区域的硅探针电生理记录相结合的方法(图1)。我们使用了表达嵌合VSFP Butterfly Y/R(chiVSFP Butterfly)转基因的小鼠,该基因在tetO启动子下表达。通过与CaMK2A-tTA小鼠杂交,chiVSFP Butterfly在新皮层和海马体锥体神经元中表达。通过将Ciona intestinalis电压感应域的一部分替换为快速激活和失活的Kv3.1钾通道的同源部分,chiVSFP Butterfly从早期的VSFPs发展而来。这种修饰加速了GEVI的响应动力学,chiVSFP Butterfly能够跟随高达200 Hz的膜电压振荡。与单色GEVIs不同,chiVSFP Butterfly在两个荧光波段中报告膜电压变化,具有相反的荧光变化。这使得比率测量成为可能,从而便于校正单色(单波长)GEVI成像中固有的血流动力学和pH相关的干扰。
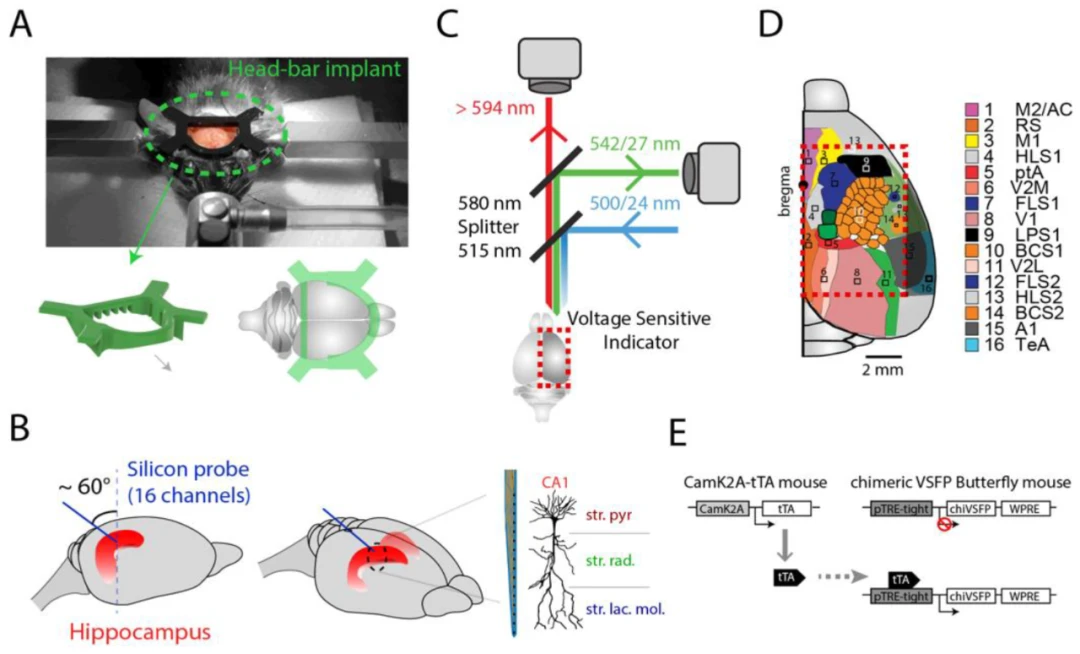
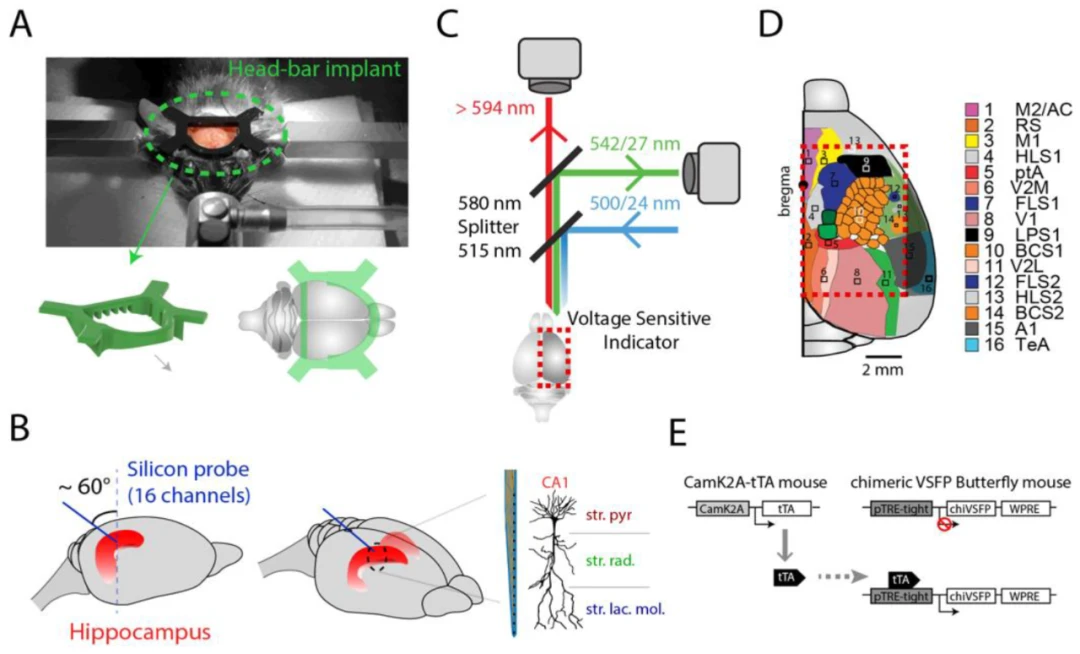
图1. 结合宽场皮层成像和海马体电生理记录的植入程序。(A)在立体定位装置上对小鼠进行头部固定杆植入。(B)将高密度硅探针(16通道)植入海马体的示意图。探针的接触点间距为50 μm,可以覆盖CA1的所有层。注意,植入物位于皮层的后部,靠近人字缝,从而使整个皮层区域在光学成像视野中不受阻碍。(C)用于宽场皮层光学电压成像的双发射宏观镜。从浅表皮层组织记录的电压指示剂的荧光发射由两个同步相机记录,随后用于比率分析。(D)宏观镜可以同时成像的多个区域的拓扑图(拓扑图像经许可改编自参考文献)。(E)使用的GEVI转基因小鼠系的示意图;CaMK2A启动子控制的嵌合VSFP Butterfly(chiVSFP)在锥体细胞中的表达(CaMK2A-tTA;tetO-chiVSFP)。
首先,我们记录了同一小鼠的两种生命状态:体内(静息状态)和体外(终末安乐死后)以比较信噪比。正如预期的那样,在体外条件下,新皮层没有显示任何活动,而在活体大脑中观察到新皮层的自发波动。同样,在体内状态下,与皮层电压成像同步的海马体电生理记录中,可以观察到海马体锥体层中的自发尖波涟漪(SWR)事件,这些事件伴随着皮层瞬态活动(图2A)。在体外状态下,海马体活动缺失,正如预期的那样。
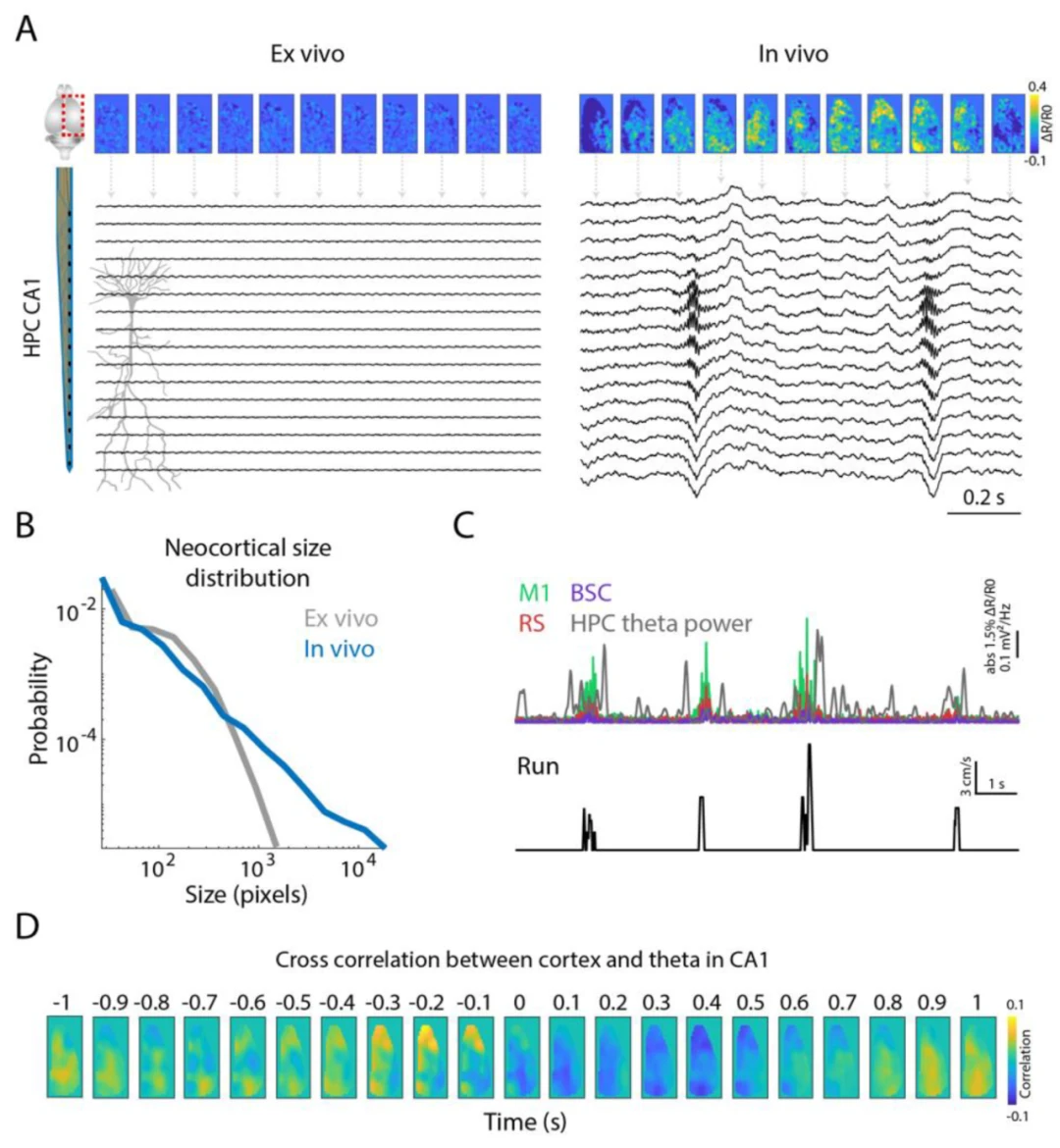
图2. 信号质量。(A)动物在体外和体内状态下的皮层电压成像和海马体CA1电生理记录示例。(B)基于活动波动空间范围(大小)的新皮层瞬态概率分布。正如预期的那样,体内状态下的瞬态遵循幂律分布。体外制备中也存在较小的瞬态,可能是由于噪声。(C)同时记录的绝对ΔR/R皮层活动和海马体θ功率在活跃(跑步期间)和安静行为期间的变化。注意,在跑步期间,动物表现出更高的皮层电压信号调制,这与海马体θ功率的增加相似。标签定义为初级运动皮层(M1)、压后皮层(RS)、桶状皮层(BSC)和海马体(HPC)θ功率。(D)皮层像素与海马体锥体层θ功率之间的互相关。
活跃行为状态下的运动对皮层范围的神经活动产生了强烈的调制。在啮齿动物海马体中,θ频带(5-10 Hz)是行走、跑步和嗅探等活跃状态的标志。与之前的研究一致,我们发现多个脑区的皮层活动在跑步机跑步期间增加。有趣的是,与海马体θ共存的主要运动皮层(M1)的皮层活动似乎与跑步更紧密相关。当将海马体θ功率与皮层活动相关联时,我们还发现M1活动在海马体θ振荡增加之前。
2.2 视觉信息整合过程中的海马体层特异性信号
在啮齿动物中,眼动也与空间导航过程相关。视觉信息对于空间导航至关重要,瞬时视觉输入不断更新自我中心和非自我中心的空间信息,而注视方向是该计算的重要输入。在这里,我们展示了刺激诱发的电压活动从视觉皮层传播到更高级的皮层区域,如压后皮层、扣带皮层和顶叶皮层(图3A,B)。此外,我们展示了将眼动追踪与海马体探针记录相结合,可以研究与眼跳运动相关的海马体层特异性活动。有趣的是,我们特别观察到在眼跳运动后,海马体CA1的分子层中出现γ频带活动。此外,这种调制在不同层之间似乎有所不同,这表明眼跳运动与海马体内的局部电路相关。最后,当将分子层中的γ频带活动与成像的皮层活动相关联时,我们观察到一组从内侧到外侧区域(顶叶、初级和次级外侧视觉皮层;初级听觉皮层)的活动先于海马体CA1中的中γ振荡。在中γ振荡期间,这种相关性似乎集中在更内侧的区域(初级和次级运动、感觉和压后皮层),这可能与动物的运动有关。
图3. 视觉信息传播到皮层联合区域和海马体。(A)视觉刺激范式的示意图。(B)特定视觉刺激模式(45°)期间的平均皮层电压活动曲线(N = 50次试验)。(C)使用DeepLabCut检测瞳孔大小的面部相机(左图)。眼跳运动期间海马体硅探针的平均局部场电位(右图)。频谱图显示了眼跳运动期间三个通道(分别位于锥体层、辐射层和分子层)的频谱平均值。(D)每个皮层像素与分子层中γ功率的互相关。
2.3 海马体神经元与皮层模块的相互关系
海马体具有多个传出和传入投射,连接包括皮层在内的不同脑区,海马体-皮层通信对于将短期皮层表征存档为长期海马体存储的记忆痕迹等过程至关重要。结合皮层电压成像与海马体硅探针记录,为在体内研究这种连接提供了独特的机会。在这里,我们使用16通道硅探针检测海马体CA1活动(图4A),在头部固定的清醒动物中进行皮层电压成像。我们检测到多个具有不同波形的神经元。接下来,我们计算了与海马体神经元放电活动相关的新皮层活动的尖峰触发平均值,并观察到与CA1锥体层不同神经元放电活动相关的不同皮层图。我们展示了这种组合方法提供了详细研究海马体-皮层关联的潜力。
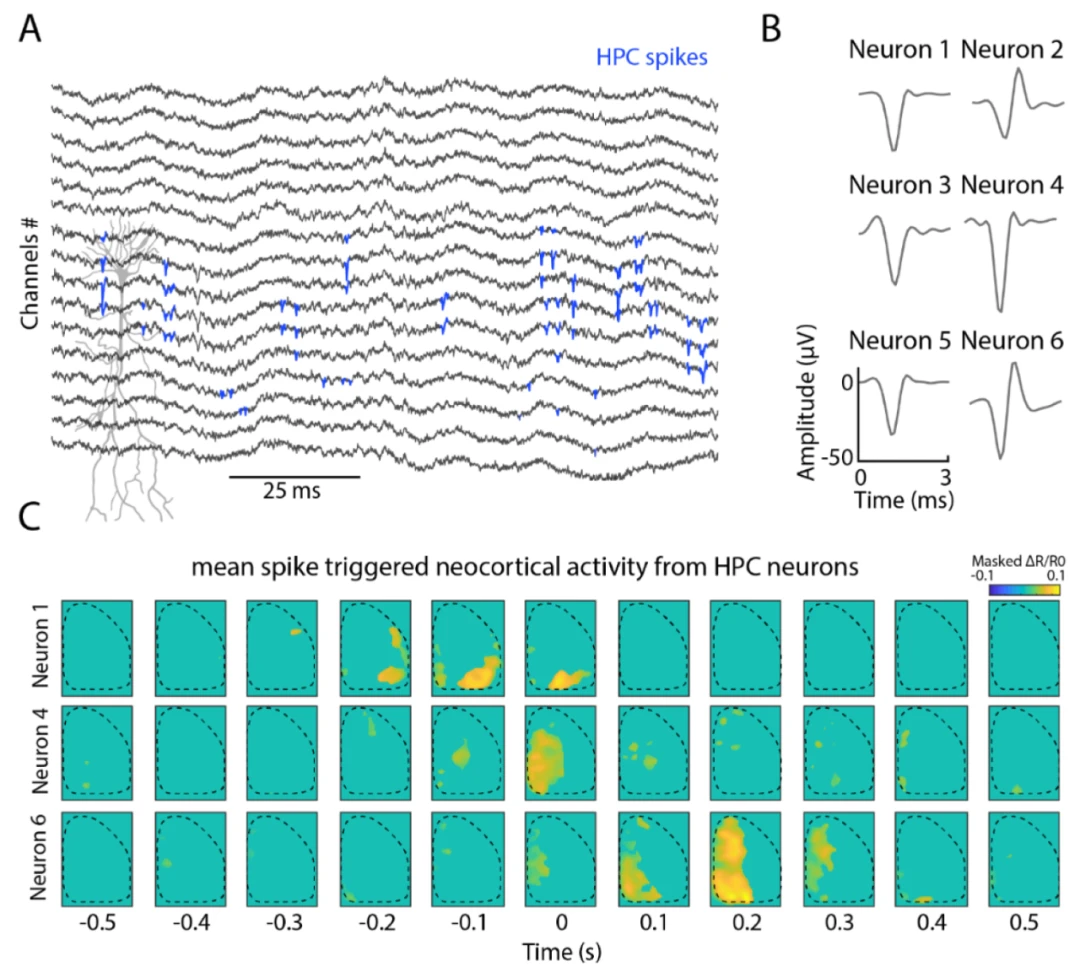
图4. 结合皮层电压成像和海马体电生理记录揭示了不同的活动模式类型。(A)海马体CA1中硅探针的局部场电位。蓝色表示使用kilosort 2.0检测到的假定尖峰。(B)在CA1锥体层中检测到的6个示例神经元的波形。(C)海马体中3个神经元尖峰活动周围的新皮层激活。图被信号平均值加5个标准差的平均值掩盖(每个检测到的神经元N = 200次平均随机尖峰)。
2.4 皮层微刺激与海马体电生理
接下来,我们展示了在体内观察皮层电压活动和对皮层微刺激的海马体电生理活动的技术可行性。为此,我们分别以不同的刺激强度(20、50、100和200 µA)电刺激压后皮层(RS)和体感皮层(SS),并监测这两个皮层结构如何与皮层半球的其余部分以及不同频率的海马体振荡相互作用。
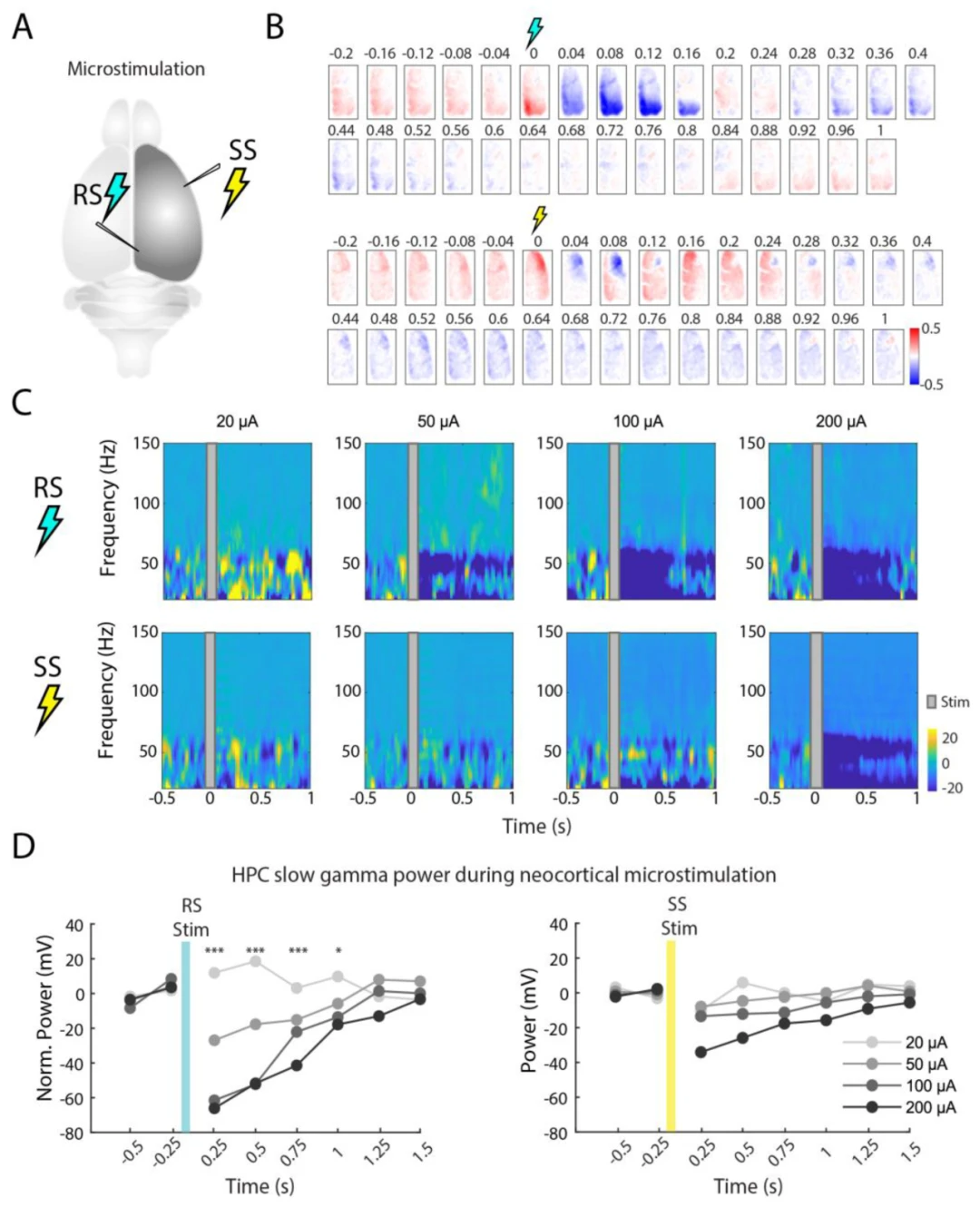
图5. 皮层微刺激对皮层和海马体活动的影响。(A)压后皮层(RS)和体感皮层(SS)微刺激区域的示意图。(B)RS和SS刺激后的皮层电压活动模式(10 ms刺激;200 µA;50次刺激事件的平均值)。注意,在短暂的增加活动期后,随后的1秒内持续抑制。(C)在20、50、100和200 µA刺激下,海马体CA1的频谱图。(D)在20、50、100和200 µA刺激下,γ(20-50 Hz)功率的基线归一化(* p < 0.05,*** p < 0.0005,重复测量ANOVA,每个条件N = 50次刺激)。
在200 µA的RS或SS刺激后,我们观察到刺激区域在皮层成像数据中显示出瞬时活动爆发,随后是持续约1秒的扩散超极化。观察不同强度(20、50、100和200 µA)刺激对海马体频谱活动的影响,我们发现RS刺激(抑制)对海马体LFP(主要在5-40 Hz频率之间)产生了干扰。这种海马体振荡干扰与RS区域中诱导的电流大小(以及随后的RS抑制)成正比。相反,SS刺激在海马体LFP中没有显示出显著差异。结合皮层成像、皮层微刺激和海马体LFP,我们的观察表明,压后皮层与海马体CA1之间可能存在因果关系,但需要进一步研究以确定这是由于皮层输入到海马体,还是由于海马体传入在压后皮层中的反激活和抑制。
三、讨论
许多系统级研究的一个常见误区是假设特定的认知反应仅涉及特定的脑区。这是因为体内记录通常设计为仅针对少数区域,而忽略了大脑其他部分的贡献。在这里,我们主张宽场电压成像可以成为在清醒、行为动物中同时观察多个皮层区域的有价值的方法。与其他脑活动代理指标(如钙和谷氨酸)相比,电压活动具有更快的
时间分辨率,类似于电生理学,这使得其能够与电生理数据结合进行因果分析。
重要的是要强调,电生理学与皮层电压成像的结合是灵活的,可以根据不同的科学问题进行调整。在这里,我们展示了将皮层电压成像与海马体电生理学结合的可行性,以开始揭示与视觉处理相关的海马体-皮层关系。
视觉信息在空间导航中起着重要作用。特别是,识别地标和边界是空间认知所需的过程,这些过程需要来自初级视觉区域的信息传递到大脑组织层次中更高级的区域,如海马体,在那里可能生成环境的认知表征。然而,视觉信号如何调节海马体中的空间表征以及信息如何传播到海马体仍然未知。为了研究这一点,我们主张宽场皮层电压记录可能提供有关视觉整合机制的重要细节。为了实现这一目标,我们在此展示了视觉皮层的活动如何传播到其他相互连接的高级皮层区域,如海马体、压后皮层、扣带皮层和顶叶皮层。此外,我们观察到眼跳运动主要与海马体CA1分子层中的中γ活动相关。将这种海马体γ活动与皮层活动相关联,我们发现一组皮层区域(其中包括视觉皮层)的活动先于海马体中的中γ振荡。因此,通过我们的技术组合,我们能够展示清醒状态下眼-海马体-皮层之间的新型相互作用。
在层次上,海马体被视为记忆过程中的重要中枢区域。它由多个散布在整个大脑中的传出和传入投射组成,其中包括皮层。经典的假设认为,海马体保留短期记忆并将其索引到皮层以形成长期记忆,这突显了海马体-皮层通信的重要性。在此过程之前,短期记忆从感觉信息的编码及其适应海马体中产生。从这个意义上说,海马体-皮层相互作用产生的尖峰调制可以解决这一计算问题。在这里,我们展示了海马体中的活动可以具有与皮层活动不同的时间和空间动态。在广泛的新皮层水平上,先前已经观察到了记忆的重新激活。结合我们的结果,我们可以推测,关于海马体事件(如重放或细胞组装的形成)的信息被传递到特定的皮层回路。
我们还展示了在压后皮层或体感皮层微刺激期间,皮层电压活动与海马体LFP的结合。我们的结果表明,海马体中的γ振荡被压后皮层刺激引起的抑制所抑制,并且这种抑制与压后皮层中的电流强度相关。之前我们报告了压后皮层活动在睡眠期间与CA1中的γ振荡相互作用,并且这种相互作用具有从皮层到海马体的时间关系。这与我们在此的结果一起表明,压后皮层-CA1通信对海马体LFP有直接影响。
总之,海马体-皮层关系在神经科学中被认为在记忆巩固机制中起着基础性作用。尽管如此,对这两个结构之间基本相互作用的研究仍存在功能空白,部分原因是技术限制。在未来几年,皮层宽场电压成像可能有助于理解不同行为状态下的层次性新皮层过程。结合海马体LFP,这种方法有潜力阐明记忆巩固等认知过程的基本电路。
四、材料与方法
4.1 动物
我们使用了CaMK2A-tTA;tetO-chiVSFP转基因小鼠(3至6个月大,体重25-35克)。该转基因小鼠系在所有皮层层的锥体神经元中特异性表达GEVI。宏观荧光成像将光学访问限制在浅表皮层。动物在12小时光照/黑暗周期下饲养,自由饮水进食。
4.2 结合宽场成像与海马体电生理的手术程序
为了记录皮层光学电压成像与海马体电生理(N = 4只小鼠),我们进行了手术以暴露头皮并慢性植入16通道线性探针(Atlas Neuroengineering,比利时,记录点间距50 μm)到海马体中。手术过程中,动物被麻醉并放置在立体定位框架中,通过鼻罩输送0.5-1.5%的异氟烷。在整个手术过程中,通过加热垫维持体温在37°C左右,呼吸频率为0.5-1 Hz。然后,在切口区域皮下注射2%利多卡因(50 μL)。我们依次暴露了大面积的头皮,并用手术钻削薄整个右半球的颅骨以减少光散射。此外,在左半球颅骨中植入两颗螺钉以提高头板的机械稳定性;第三颗螺钉也植入同一半球小脑的浅表区域,用作接地和参考。对于海马体电生理记录,将16通道线性探针慢性植入右半球(成像皮层的同侧)。探针最初放置在+2.4 mm ML和-4.3 mm AP,角度为60度,然后降低并固定在约2.2 mm深度。最后,小心地将头板放置并用丙烯酸水泥固定在颅骨上。
4.3 结合宽场电压成像与海马体电生理
我们在术后恢复后对小鼠进行同步皮层电压成像和海马体LFP记录。电压成像的光学配置如前所述,使用宽场双发射宏观镜。荧光激发由高功率卤素灯提供,我们同时使用两个同步sCMOS PCO edge 4.2相机测量mKate2(GEVI FRET受体)和mCitrine(GEVI FRET供体)的荧光发射。光学配置包括:mCitrine激发500/24和发射FF01-542/27,mKate2发射BLP01-594R-25,分束器515LP和580LP。图像系列以50 Hz帧率从右半球皮层获取,分辨率为375×213像素(约60像素/mm),采样深度为12位,每次记录10分钟。
为了计算光学测量的电压活动,计算了mCitrine和mKate2荧光发射的增益均衡比率(ΔR/R),然后归一化为ΔR/R = (R(t) - Rmean)/Rmean;Rmean为会话平均比率。我们隔离并减去了心跳频率成分,并应用了>0.5 Hz的高通滤波器以去除残余的血流动力学信号。为了量化皮层电压波动,我们使用类似于Scott等人的方法检测活动瞬态,其中每个像素的激活状态必须超过定义为总激活帧平均值的阈值。
与皮层电压成像同步,使用16通道头戴式放大器(Intan Technologies,RHD2132)和Open Ephys系统获取海马体CA1的局部场电位(LFP)。原始信号以0.1-7500 Hz滤波并以20 kHz采样,然后降采样至1 kHz进行分析。我们使用kilosort 2.0进行尖峰排序。θ功率通过滤波后的LFP信号在5-10 Hz之间的包络计算。
4.4 视觉刺激呈现
视觉刺激呈现在距离小鼠左眼10厘米的显示器上。我们呈现了50个45度的光栅,每次持续1秒,间隔随机在1到2秒之间。
4.5 眼动追踪与瞳孔检测
为了检测记录期间小鼠眼球的眼跳运动,我们使用固定相机(Basler aca1920-150 um,f = 20 mm镜头)对准头部固定小鼠的面部,并使用红外照明以20 Hz获取图像。我们使用DeepLabCut标记每个行为成像帧中检测到的瞳孔大小。通过瞳孔中心的高运动,我们通过阈值化眼球速度获得了眼跳时间。
4.6 皮层微刺激与结合皮层电压成像和海马体电生理
对于皮层微刺激,我们使用了两根钨电极(直径50 μm)的束,电极尖端垂直距离约为250 μm。植入手术程序如上所述,增加了在头板固定前将两根刺激电极束慢性植入压后皮层(RS)和体感皮层(SS)的步骤。对于RS植入,我们首先在右半球颅骨上进行了小颅骨切开术,坐标为+0.5 mm(ML)和-2.5 mm(AP),并将束电极植入+0.5 mm(DV)并向中线倾斜45度。然后在右半球坐标为+3 mm(ML)和+1 mm(AP)的SS皮层上方进行了小颅骨切开术。束电极在这种情况下也植入+0.5 mm(DV),但向脑外侧倾斜30度。
术后恢复10天后,将清醒动物放入记录装置中进行皮层刺激与皮层电压成像和海马体电生理的结合。我们使用Arduino UNO发送50个10毫秒的方波脉冲,刺激间隔随机在1.5-2.5秒之间,控制刺激隔离器(World Precision A365),并与Open Ephys同步。刺激强度为20、50、100和200 µA。
东莞富临医疗科技有限公司是Open Ephys 和 Intan Technologies 在亚洲的代理商,为亚洲客户提供“技术服务”与“电生理产品”
邮:li@fulinsujiao.com
公司地址:广东省东莞市樟木头镇塑金国际1号楼810

