
不同物种的竞争也是网络动物圈绕不开的一个话题,而物种竞争动态有一个经典模型——“洛特卡-沃尔泰勒(Lotka–Volterra)种间竞争模型”, 这里我将结合自己对基础生态学的一点点浅薄学习与对另一位up专栏的理解来简单讲一下这个生态模型。

在了解“洛特卡-沃尔泰勒种间竞争模型”之前,我们需要先来解读一下几个种群增长模型。
首先,先看一下与密度无关的种群增长模型,这个“与密度无关“意味着空间和资源无限,种群可以以类似实验室不受限制条件下增长(即以内禀增长率增长的种群)。这种种群增长情况又分为两类:离散型和连续型。
1 离散型意味着种群各个时代不重叠(如一岁一枯荣的一年生植物与昆虫),种群增长不连续,下一代在繁殖时上一代已经无了。
若Nt为t世代种群的大小(Nt+1则为t+1世代种群大小),λ为增长率,则种群离散增长模型可表示为:
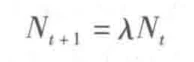
若种群每年都保持λ的增长率,则为:
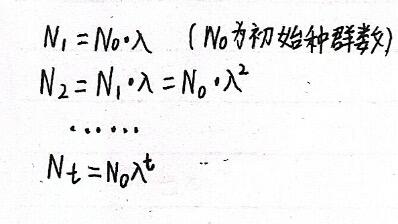
为了简化运算,将这个指数方程两边分别取对数,可得:
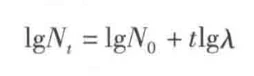
所以以lgNt对t作图可得这张图(当然图里面lgλ>0且A>1,种群咋样要看λ):
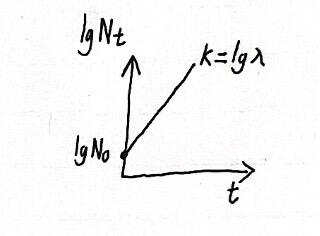
2 连续型意味着时代间有重叠,也考虑死亡的情况,但不考虑迁入和迁出。假定在很短的时间dt内的种群瞬时出生率为b死亡率为d,则种群瞬时增长率r=b-d。在这个模型中增长率与种群密度无关(因为种群空间资源无限)。
可得:

(别问我怎么用一阶线性微分方程通解得出这个积分式的,我数学不好)
这样一来种群增长曲线就是“J”型的了,而如果对数为纵坐标则为直线型。
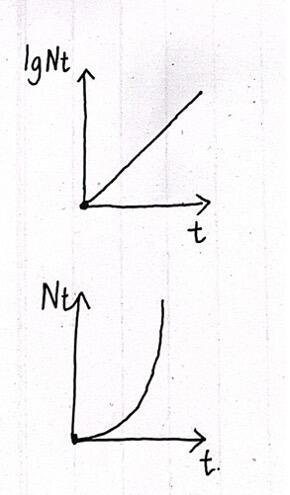
但是自然界几乎不可能有种群无限增长的情况,因为空间和资源毕竟是有限的,因此生态学家发明了逻辑斯蒂方程,这个方程可以简单看作是把上面提到的指数增长方程乘上一个密度制约因子。
具体来说,逻辑斯蒂种群连续增长方程增加了两个条件:1存在一个环境容纳量K,当种群数量Nt=K时,种群将不再增长,而dN/dt(表示单位时间内种群净增长率)也将等于0。2假设一个空间只能供K个个体生存,每一个个体利用1/K的空间,那么N个个体就利用N/K的空间,这样以来剩余可以供该物种生存的空间就只剩下(1-N/K)了(即每多一个个体,就会对增长率产生1/K的抑制)。
根据上面两个条件,种群瞬时增长率r将随着种群密度的增加而下降。种群增长曲线也从的“J”型变成“S”型。曲线上升平滑且最终会无限接近K值。
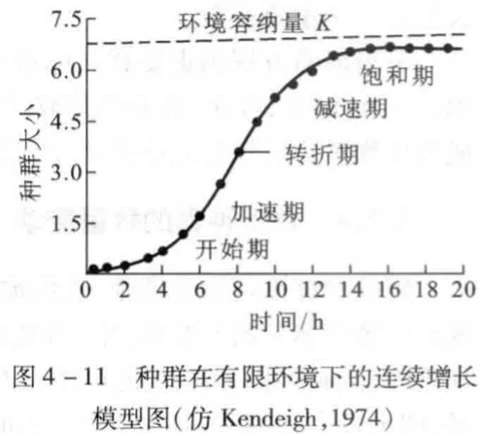
那么逻辑斯蒂方程则可表示为:

接下来看洛特卡·沃尔泰勒(Lotka–Volterra)种间竞争方程,这个方程可以视为逻辑斯蒂方程的延伸。
按照逻辑斯蒂方程,在单独情况下,舍N1、N2分别为两个物种的种群数量,K1、K2、r1、r2分别为两个物种的环境容纳量和种群增长率,则可列出:
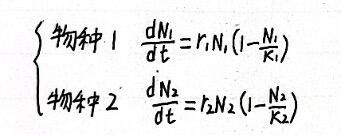
当两个物种共同利用同一生存空间时,需要引入竞争系数α和β,相当于一个N2等同于α个N1的竞争效果,换算成N1就是α*N2。β同理。这样,洛特卡·沃尔泰勒种间竞争方程可列为:
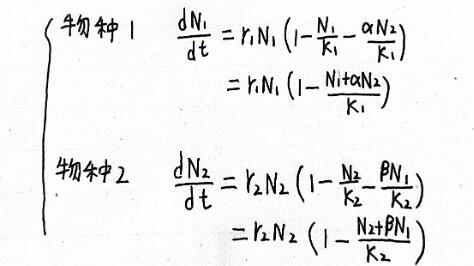
(很多人可能产生这样的疑问,为什么不是α*N1?我们可以这样去理解,比如N1和N2都各有一只,但N2所占空间相当于10个N1所占的空间,而N2代表的是数量,所以就要用N2乘上10。所以上述分母上α*N2和β*N1也是同理)
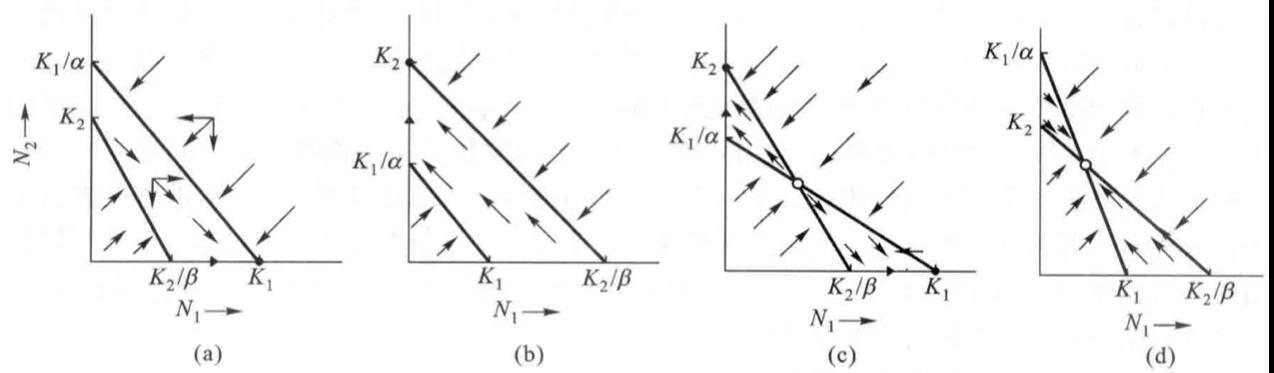
有了这个式子,我们可以从物种1的角度去试想两种极端的情况:1.N1占据全部生存空间,N2消失,则N1=K1,N2=0;2.N2占据全部生存空间,则N1=0,N2=K1/α。将这两个端点从坐标轴连起来,则得到N1零增长线。同理也能得到N2零增长线,箭头代表了不同种群数量基数下种群数量的变化趋势。
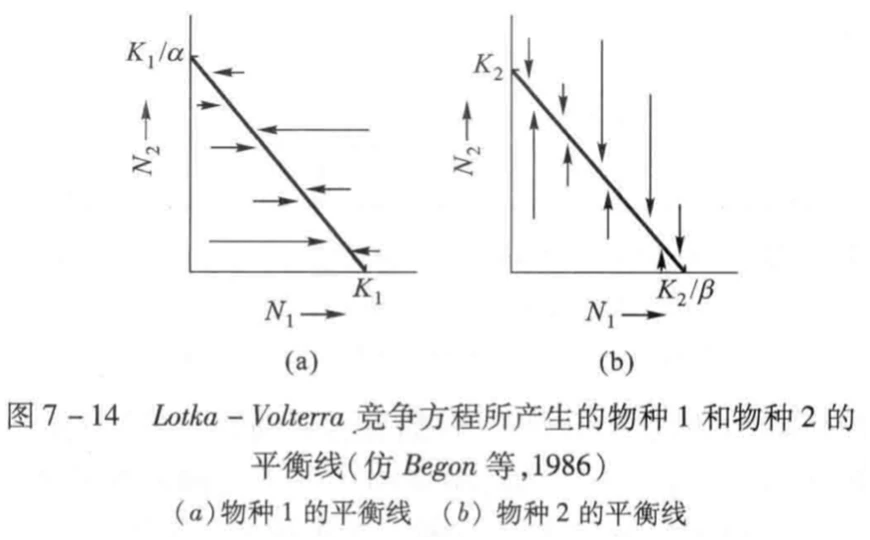
如果将这两个图叠在一起,则可得到四种不同的竞争结果,而该结果取决于K1、K2、K1/α和K2/β的相对大小(即环境容纳量和物种的竞争系数)。
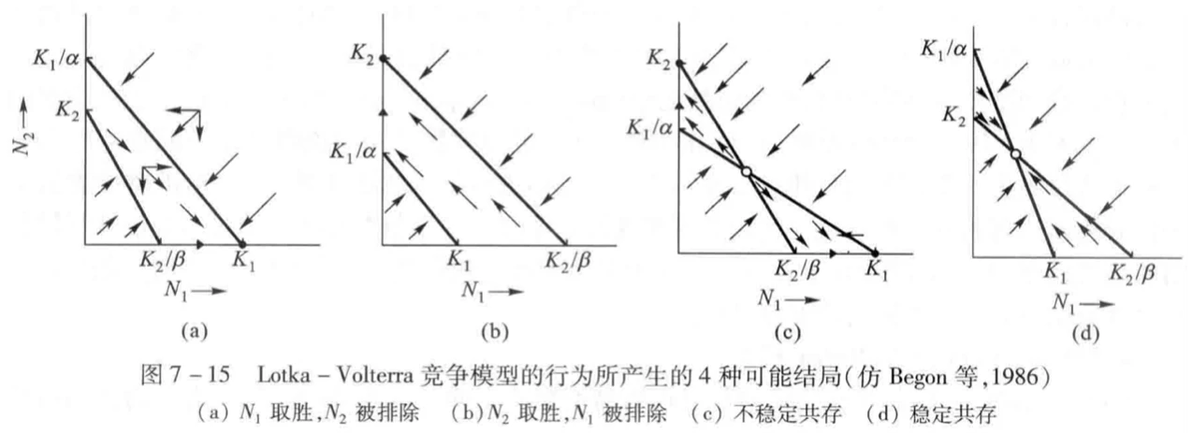
(1)若K1>K2/β,K2<K1/α,则N1在竞争中获胜,N2被完全排挤掉(这个可以在图上用“向量”来理解,合向量箭头最终指向N1=K1,N2=0的区域)。
(2)若K2>K1/α,K1<K2/β,则N2在竞争中获胜,N1被完全排挤掉。
(3)若K1> K2/β,K2>K1/α,两条线相交,出现平衡点,但不稳定(合向量箭头指向两边)。
(4)若K1<K2/β,K2< K1/α,两条线相交,出现平衡点,平衡点稳定(合向量箭头指向平衡点)。
根据这个种间竞争模型,1/K1和1/K2可作为两个物种各自的种内斗争强度指标,那么环境容纳量越大,种内斗争(1/K)就相对越小。
同理,β/K2可作为物种1对物种2的种间竞争强度,α/K1可作为物种2对物种1的种间竞争强度。
由此可见,竞争的结果取决于种间竞争强度和种内斗争的相对大小。
(1)1/K1<β/K2,1/K2>α/K1;物种1内斗弱外斗强,物种2内斗强外斗弱,物种1将取代物种2,即图(a)
(2)1/K1>β/K2,1/K2<α/K1;物种1内斗强外斗弱,物种2内斗弱外斗强,物种2将取代物种1,即图(b)
(3)1/K1<β/K2,1/K2<α/K1;俩物种都是内斗弱外斗强,谁都有可能取代对方,所以呈现图(c)的不稳定平衡状态
(4)1/K1>β/K2,1/K2>α/K1;俩物种都是内斗强外斗弱,谁取代不了对方,所以呈现图(d)的稳定平衡状态
根据上面提到过的逻辑斯蒂方程中的密度制约因子的来由并结合@动物志-剑齿猫的文章《抛开数量谈竞争力是耍流氓,用模型推演虎豹、虎狼、狼猞猁的竞争》,我们可知环境容纳量K不仅与环境资源有关,更大程度上也与动物的种内斗争有关(如狼群会建立排外的领地,在种群较为饱和的地区,不同狼群之间的打斗是其自然死亡事件发生的主因),而K值越小则往往反映出种内斗争的程度,即种群只能容纳较少个体。
当1/K1<β/K2即K1>K2/β时,物种1的内斗较小,能够达到排挤掉物种2的数量。
出了环境容纳量K以外,竞争系数α和β也是种间竞争结果的重要影响因素,而在@动物志-剑齿猫的文章最后提到,竞争系数α和β也与环境有很大关系,因为环境可以影响不同物种之间资源利用性竞争和干涉性竞争的强度(如环境资源的丰富度可以使两个物种的生态位在一定程度上分开,或者适宜的环境有可能使某一物种更容易逃离其他物种的攻击。
而网络上的斗兽话题可能喜欢强调某物种战斗力强且凶猛好斗,这样的动物当然在与其他动物的打斗中更有可能驱赶甚至杀死对方,但也意味着它们的种内斗争也可能更加激烈,这样的话我们将物种1降低的环境容纳量K带入上面种间竞争强度与种内斗争强度指标可知,物种1的内斗强度与物种2对物种1的外斗强度也会增加,所以根据上述理论,战斗力强且凶猛好斗对种间斗争结果的影响是不确定的,也就是说并不是说战斗力强且凶猛好斗就一定能够使这种动物一定能在竞争中取胜。
@动物志-剑齿猫的专栏:

