

1.长薄鳅(Leptobotia elongata)的分类地位
鲤形目(Cypriniformes)鳅科下分三个亚科:花鳅亚科,条鳅亚科和沙鳅亚科。长薄鳅所属的薄鳅属隶属于沙鳅亚科。

长薄鳅,图片来自网络,侵删
2.长薄鳅(Leptobotia elongata)简要形态特征
体长,侧 扁,尾柄高而粗壮。头侧扁而尖 ,头长大于体高 。吻 圆 钝 而 短 , 口 较 大 , 亚 位 , 口裂呈马 蹄 形 。上下唇肥厚 ; 唇稱与颂分离 ,颐下无钮 状 突 起 。 须3对 , 吻 须2对 , 口角须1对 。 眼 很 小 , 眼 缘 有1根 光滑的硬刺 , 末 端 超 过 眼 后 缘 。 鼻 孔 靠近 眼 前 缘 , 前 鼻 孔呈 管 状 , 后 鼻 孔 较 大 , 前 后 鼻孔 之 间 有1分离 的 皮 稱 。 鲍 孔较 小 , 鲍膜 在 胸 鰭 基 部 前 缘 与 峡部侧 上 方 连 接 。 背鳍和 臀鳍 均 短 小 , 没 有硬 刺 ; 背鳍 位 于 体 的 后 半部 ; 胸 、 腹 鳍 短 , 胸 鳍 基 部 具 有1个长 形 的 皮 裙 ; 尾 轄 深叉 状 。 麟 极 细 小 。 侧 线 完 全 。头 部 背面具 有 不 规 则 的 深褐色 花 纹 , 头 部 侧 面及觸 盖 部 位 为 黄 褐色 ,身 体 浅 灰 掲色 。较小个体有 6-7条很 宽的深 褐 色 横 纹 , 大 个体 则 呈 不 规 则的 斑纹 。腹部 为 淡 黄 褐 色 。背 鳍基 部 及靠 边缘 的地方 , 有 两 列 深 褐色 的 斑 纹 ,背 轄带有黄 褐 色 泽 。 胸轄及腹 鐘 呈 澄 黄 色 , 并 有褐色斑 点 。 臀 轄 有2列褐色的斑 纹 ; 尾 鰭浅黄褐 色 , 有3-4条褐色条纹 。
3.长薄鳅(Leptobotia elongata)的大致分布
长薄鳅是我国特有的种类,在我国分布较广,遍布长江中上游干流及其支流,还在云南的元江中也有捕获。干流中宜宾-重庆段的长薄鳅产量最高,支流中嘉陵江和雅砻江产量较高。
4.长薄鳅(Leptobotia elongata)遗传多样性
近年来,由于过度捕捞及长江中上游水电站建设导致的水域分割及水流条件改变等原因,长薄鳅野生资源逐年减少,因此开展其遗传多样性研究对保护和利用该种鱼类具有重要意义 。遗传多样性衡量常用单倍型多样性(Hd)和核苷酸多样性(π)。赵刚等采用线粒体DNA控制区D-LOOP分子标记对泸州、南溪、柏溪、攀枝花和重庆5个群体的长薄鳅控制区遗传多样性进行了研究,结果发现 Hd=0.916;π=0.004 50;Liu等同样对以上5个群体的长薄鳅控制区遗传多样性进行了研究,结果发现Hd=0.952;π=0.004 5,两组研究结果差异显著。田辉伍等对长薄鳅控制区遗传多样性进行了研究,结合历史结果得出结论:长薄鳅种群或种群内部分地理群体在较近时间内发生过种群扩张事件;并对赵刚等和Liu等的研究进行分析,认为两组结果差异显著的原因可能是赵刚等的样本群体不完整和样本缺失导致单倍型的检出率低 。Liu等 和连灏等通过对微卫星位点的比较研究,评估了长江上游8个地点(新市、攀枝花、合川、赤水、乐山、宜宾、江津和三峡库区)的长薄鳅遗传多样性,并分析了种群结构,发现野生长薄鳅种群的遗传多样性仍保持在较高水平。Liu等利用线粒体DNA控制区和核微卫星 DNA 标记,研究了金沙江下游建坝前 (2008年)和建坝后(2014—2015年)12个种群的遗传多样性和种群结构,发现在金沙江下游建坝后,该区域野生长薄鳅的遗传多样性没有下降,而且大坝上下游之间有形成种群结构的趋势 (2020年)。但与同水域的红 唇 薄 鳅 (Leptobotia rubrilabris)、 中 华 沙 鳅(Botia superciliaris) [20] 相比,长薄鳅的遗传多样性指数仍处于较低水平 。Li等测序了长薄鳅全线粒体基因组序列,发现在系 统 进 化 中 , 长 薄 鳅 与 松 花 江 薄 鳅 (Leptobotia mantschurica) 同 属 薄 鳅 属 , 薄 鳅 属 和 沙 鳅 属(Chromobotia) 构 成 姐 妹 群。Leptobotia 和Chromobotia 为 一 枝 , 组 成 沙 鳅 亚 科 (Botiinae);Acantopsis、 Cobitis、 Pangio、 Niwaella、 Koreocobitis、Misgurnus 属为一枝,组成鳅亚科 (Cobitinae)。两枝共同构成长薄鳅的进化树。据统计,长薄鳅遗传研究中采用微卫星标记方法的占比 58.30%,采用线粒体DNA的研究占比41.70%。
5.长薄鳅(Leptobotia elongata)的生活习性
长薄鳅为广温性鱼类 ,喜钻泥,生活在水流较缓、溶氧含量高的河流底部,体色可以随周围环境微变,水温0~33 ℃下均可生存,最佳生长温度为23~28 ℃,有较强的耐饥饿能力,无需停食越冬,冬季亦可在室外浅水中生存。长薄鳅的热耐受能力较差,水温超过33 ℃即可致死 。鱼类的昼夜间耗氧率变化规律反映了鱼类的活动周期。邹桂伟等应用测定流水中溶氧量的方法测定了长薄鳅的耗氧率,发现长薄鳅的耗氧率具有昼夜节律性且受温度影响 。丁少波等对大渡河下游典型鱼类的游泳能力进行了测试,发现在研究的 6种鱼类 (胭脂鱼、长薄鳅、长鳍吻鮈、异鳔鳅鮀、唇鱼骨、四川白甲鱼)中,长薄鳅的游泳能力位居前列 。
6.长薄鳅(Leptobotia elongata)的周年生长特性
不同种类的鱼类在不同的生长阶段具有不同的生长强度,即鱼类生长具有阶段性 ,且同种鱼类在周年的生长过程中在不同的养殖季节、不同的养殖水温环境下也会表现出不同的生长特性。从本试验的数据分析中可知 5 -8 月份水温上升时期,即水温月均值范围 21. 9 ~ 25. 49 ℃期间,是长薄鳅 1 龄幼鱼的快速生长阶段,同时试验数据也表明在一定的温度范围之内,随着温度的升高,生长也在加快。这与谢永德关于菊黄东方鲀( Takifugu flavidus) 1龄幼鱼生长特性、鲁宏申关于达氏鳇( Huso dauricus) 1 龄幼鱼生长特性、牟振波关于细鳞鱼( Brachymystax lenok) 生长特性等的研究结果相似。
7.长薄鳅(Leptobotia elongata)所处水温与生长关系
水温是影响鱼类生长的重要因素,适温条件对鱼类的生长具有促进作用,不同鱼类所适应的水温生态幅也具有差异性,温度可以直接影响到鱼类的生长、发育、新陈代谢等生命活动过程 。通过分析长薄鳅周年生长数据和体长生长、体重生长、特定生长率与水温的关系可知,2017 年 5 - 8 月份水温上升阶段是长薄鳅快速生长的时间段,这期间的水温月均值变化范围在 21. 90 ~25. 49 ℃,即水温月均值在 21. 90 ~25. 49 ℃的上升期间,长薄鳅的生长迅速,是周年生长最快的阶段,当水温月均值开始下降并低于21. 90 ℃时,长薄鳅的生长速度下降,生长变缓。这与李文龙关于温度对大菱鲆( Scophthalmus maximus) 幼鱼生长、李泽滨关于温度对公子小丑鱼( Amphiprion ocellaris) 幼鱼生长、史晋绒关于温度对皇冠沙鳅( Chromobotia macracanthus) 幼鱼生长的研究结果一致,即不同种类的鱼所适应的温度范围不同,在不同的水温环境中表现出不同的生长特性。
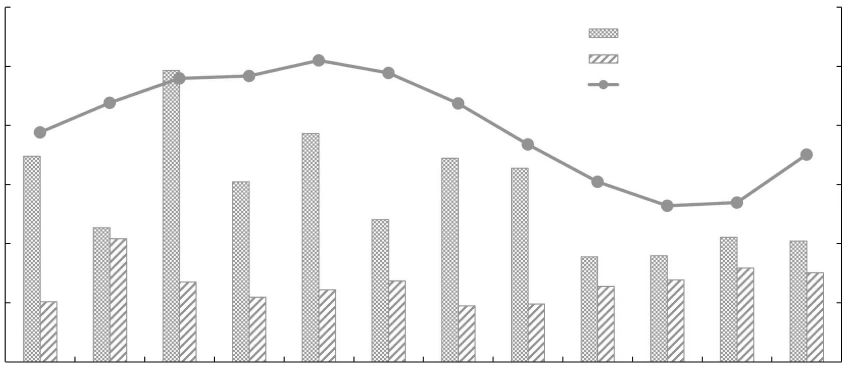
长薄鳅肥满度变化与水温的关系
8.长薄鳅(Leptobotia elongata)的人工繁殖(简)
张君等选用多潘立酮(DOM)、鲤鱼脑垂体(PG)和鱼用绒毛膜促性腺激素(HCG)三种激素作为催产激素 。催产采用两针注射法,雌鳅注射2次,雄鳅注射1次,剂量视发育情况 (发育好则精子遇水扩散快,发育一般则慢) 而定,为雌鱼的1/3~1/2。长薄鳅是分批次产卵 ,经两针注射后雌性亲鱼可顺利产出卵粒,但卵粒产出不全,卵巢中有大量遗存。经一次注射后雄性亲鱼可顺利产出精子,且受精率较高 。在人工繁殖中,长薄鳅野生后备亲鱼在驯养一段时间后普遍出现性腺发育停止、甚至退化的现象,给人工繁殖造成巨大的障碍。对此,梁银栓等使用0.5 mg·kg -1 的鲤鱼脑垂体(PG)防止性腺退化,短期效果较好。
9.长薄鳅(Leptobotia elongata)的胚胎与早期发育
长薄鳅的卵径(1.50~1.66 mm)大于中华沙鳅和泥鳅 (Misgurnus anguillicaudatus)等多数鳅科鱼类,卵径较大的卵子通常有更长的内源性营养期,可以有效提高仔鱼的存活率。受精卵吸水后的卵周隙(1.24 mm)也较大,大的卵周隙可以提供给胚胎较为稳定的发育内环境,保证其正常发育。长薄鳅有着独特的脱膜方式,胚体在脱膜前期完全伸直,向前冲击破膜而出 。据张运海等报道,在24 ℃水温条件下,长薄鳅仔鱼的平均出膜时间为31.0 h。黄江文等研究发现,在19.6~20.5 ℃水温条件下,仔鱼84 h后才能完全出膜 。长薄鳅胚胎发育的适宜水温为23~24 ℃。长薄鳅仔鱼鳞片和腹鳍同步发育,鳞片覆盖臀鳍处时鳍条发育完成。发育过程历时17 d,胸鳍最早形成于幼鱼开口摄食时,此时尾鳍只出现部分鳍条,背、腹、臀鳍刚出现鳍基,最后完成尾鳍分叉。
10.长薄鳅(Leptobotia elongata)幼鱼的胃排空特征
鱼类的胃排空受温度、季节、鱼体质量、代谢率、胃肠饱满度、饵料性质和生理状况等因素的影响。鱼类在长期演化过程中,形成了一系列适应各自食性类型和摄食方式的消化道结构,因此鱼类胃排空的方式也存在种间差异。目前最常见的有3种表现形式:第1种特征为摄食最初阶段排空速度最快,随着胃饱满度的下降,排空速度减慢 ;第2种特征是胃内含物重量随时间呈直线下降的趋势 ;第3种方式表现为排空速度由慢及快再减慢的过程,不同鱼类的胃排空方式是其摄食习性和对不同食物类型适应的集中表现。测量鱼类胃排空的常用方法除了胃内含物质量分析法外,还包括放射性标记法、染色法和X-射线法等。胃内含物质量分析法研究了平均体重为(9.87±0.66)g的长薄鳅的胃排空特点,其排空特征具体表现为摄食开始至摄食后8时这一阶段快速排空,之后为一个缓慢下降的排空过程。通过分析认为,长薄鳅是肉食性鱼类,主要以小鱼、甲壳类(虾类)、底栖无脊椎动物和水生昆虫幼虫、大型浮游动物等为食,长薄鳅摄食后,大型浮游动物、底栖无脊椎动物、鱼肉等物质在消化酶的作用下较快被吸收,表现出快速减少的过程,之后由于动物性食物外壳、骨骼等较难消化部分对消化酶的化学和物理屏障作用,延迟了食糜的排出时间,进而表现出缓慢的下降过程。
11.长薄鳅(Leptobotia elongata)幼鱼的胃排空时间及其对摄食的影响
鱼类的胃排空程度与其食欲恢复密切相关。一般认为,食欲恢复的极限点为胃内残留物为零或胃内含物排空达100%,当鱼类胃内含物排空达80%时,其食欲基本得到恢复。在鱼类食欲恢复的时候投喂,可提高食物转化效率,促进鱼类生长,根据排空率制定鱼类的投喂策略具有生产指导意义。用胃排空模型可以预测胃排空时间,本研究根据指数模型,在水温23℃的条件下,长薄鳅幼鱼的胃排空率为0.2004g/h,有关研究表明,在水温23℃的条件下,体质量为10.47g的俄罗斯鲟幼鱼胃排空率为0.066g/h;体重为8.52g的许氏平鲉在水温22 ℃的条件下,胃排空率为0.047g/h。和其它鱼类相比,长薄鳅的胃排空率较大,其消化食物的速度也相对较快,预测其胃排空达到50%(食欲恢复50%)所需时间为3.5h,食欲基本恢复(胃排空达到80%)时间为8 h,食欲完全恢复(胃排空达到99%)时间为23h,即理论上投喂间隔8h时长薄鳅的食欲已基本得到恢复。但在养殖中发现,在白天自然光照环境下,即使长薄鳅食欲基本得到恢复,但其摄食率仍然偏低,结合其摄食节律,摄食率升高发生在傍晚18:00以后,直至夜间达到最大,因此,综合考虑其摄食节律及胃排空特点,建议长薄鳅幼鱼的投喂时间可安排在傍晚以后,每天投喂一次。
对了,下次专栏专门写一下长薄鳅的致危因素和保护对策
参考资料:人工养殖条件下长薄鳅幼鱼周年生长特性的研究
长薄鳅生物学特征的初步研究
中国濒危动物红皮书[M]
长江上游保护区长薄鳅和红唇薄鳅种群生态及遗传结构比较研究
长薄鳅仔,稚鱼摄食节律和日摄食率研究
长薄鳅(Leptobotia elongata)野生资源分布及其种群遗传多样性研究[D]
投喂频率及投喂时间对长薄鳅仔稚鱼生长性能的影响[J]
长薄鳅幼鱼摄食节律及其胃排空模型的研究
长薄鳅生物学特性及繁殖技术研究进展
长薄鳅的性腺发育和生殖细胞的发生[D]
长薄鳅人工繁殖技术的研究[J]
