
开发高效率、多功能性、产品高纯度和序列特异性的基因编辑工具一直是生命科学领域的长期目标。RNA可编程CRISPR系统(RNA-programmable CRISPR systems)的发现是编辑哺乳动物细胞基因组的一个里程碑,引发了三类技术的创建:CRISPR相关(Cas)核酸酶(CRISPR-associated (Cas) nucleases),碱基编辑器(base editors)和先导编辑器(prime editors)。
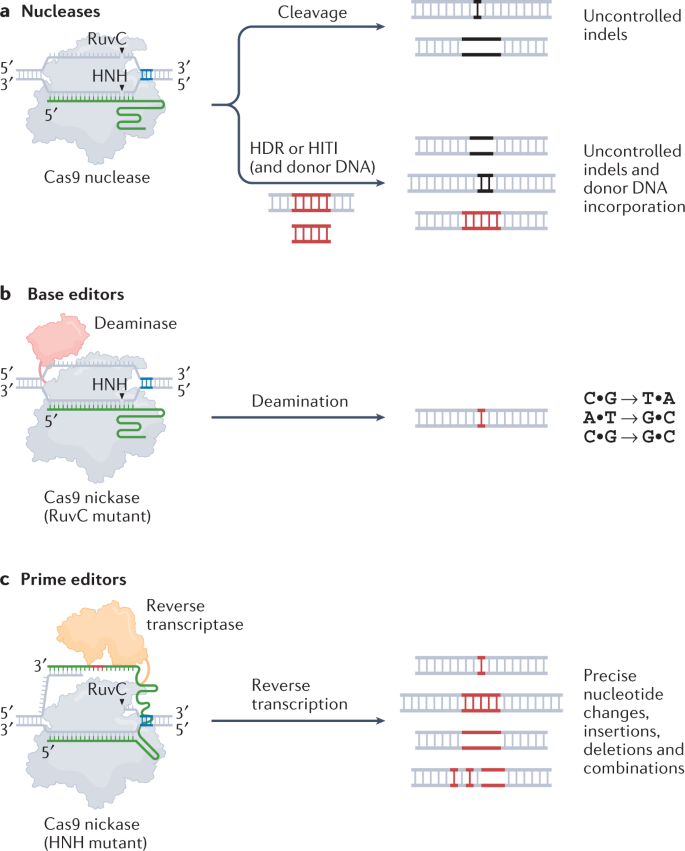
其中prime editors是同时实现多功能性、特异性和精确性的一种独特方式,不需要进行双链DNA断裂(DSB),并且能在活细胞的DNA中实现几乎任何替换(substitution)、微小插入(small insertion)和微小缺失(small deletion)。prime editing(PE)需要一个与聚合酶融合的可编程切口酶(nickase),以及能定向靶位点且以所需基因组编辑为模板的扩展向导RNA(guide RNA)。
prime editing是美籍华裔科学家刘如谦(David R.Liu)团队开发的精准基因编辑系统,近期他们在《Nature Reviews Genetics》上发表了一篇综述,全面总结了产生程序性基因组变化的主要编辑策略,强调了它们的局限性,介绍了能规避其中一些瓶颈问题的最新研究进展,并讨论了prime editing的应用和未来方向。

一、prime editing的开发和优势
Liu及其同事通过将化脓性链球菌Cas9(SpCas9)H840A切口酶的突变体,与来自莫洛尼鼠类白血病病毒(MMLV)的野生型逆转录酶(reverse transcriptase,RT)相融合,创建了最初的prime editing(PE1)。prime editing还使用一种工程化的引导RNA——prime editing guide RNA(pegRNA),它包含一个间隔序列,能将PE1引导至基因组靶位点,以及一个关键的3ʹ扩展区,用于模板化所需的编辑序列。在细胞内,prime editing–pegRNA复合物结合到和pegRNA间隔区互补的基因组靶位点上并形成R loop,其中移位的单链RNA(ssDNA)被prime editor切开一个缺口,释放一个3ʹDNA末端,可以与pegRNA延伸杂交,并引发pegRNA模板区的逆转录。通过引物延伸靶DNA链进行DNA聚合,然后生成3ʹDNA flap,包含下游基因组序列的编辑和同源物。接下来,3ʹ flap通过 flap相互转换(flap interconversion),置换基因组DNA的相邻链。切除置换的5' flap后连接剩余的缺口,形成一个异源双链,其中一条基因组链包含编辑序列。最后,DNA修复或复制将编辑序列复制到互补链,并使prime edit永久存在。
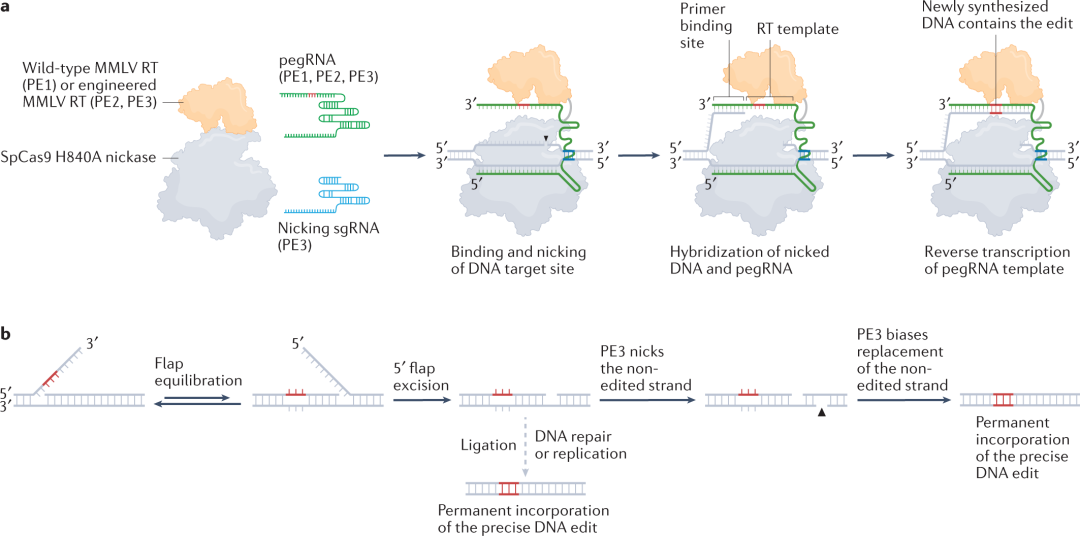
PE1可以介导人类细胞中的单碱基置换、微小插入和微小缺失,但第一代系统的效率很低,通常<5%的靶向等位基因被转化为所需序列。为了提高编辑效率,Liu及其同事向MMLV RT引入了五种突变,产生第二代editor(PE2),编辑效率提高了1.6至5.1倍。对于需要高编辑精度的应用而言,可以使用PE2系统的prime editing,并且很少生成indel副产物(通常小于编辑结果的0.5%)。
但是PE2仍依赖内源性修复过程才能将编辑从新合成的DNA链复制到互补链。之后,Liu及其同事设计了第三代editor(PE3),该系统使用额外的single-guide RNA(sgRNA)来引导prime editor酶去切割非编辑链,进一步将编辑效率提高了1.5至4.2倍。然而,PE3也诱导了一定的indel生成,这可能是由于两条DNA链上偶尔同时出现缺口,导致了DSB。使用PE3b系统,这些indel副产物可以在不损害编辑效率的情况下被消除,其中互补链缺口对编辑序列是特异的,以最小化两个基因组链上的重合缺口。PE3和PE3b支持多种人类细胞类型中高度通用的靶向替换、插入和缺失,包括初级、非分裂的小鼠皮层神经元。
与Cas核酸酶和碱基编辑器相比,prime editors为基因组编辑提供了独特优势。由于所需的编辑是在pegRNA模板内编程的,因此可以高精度地安装多种编辑类型。而且prime可以改变初始PE介导切口的远端碱基(至少33 bp),因此,与在编辑位点附近需要PAM序列的碱基编辑或Cas核酸酶介导的同源重组修复(HDR)相比,prime editing受到的限制较小。此外,prime editing很少在非目标基因组位点诱导DNA改变。与Cas核酸酶和碱基编辑器不同,prime editing需要互补碱基配对的三个检查点才能进行高效编辑。
二、prime editing的限制和改进
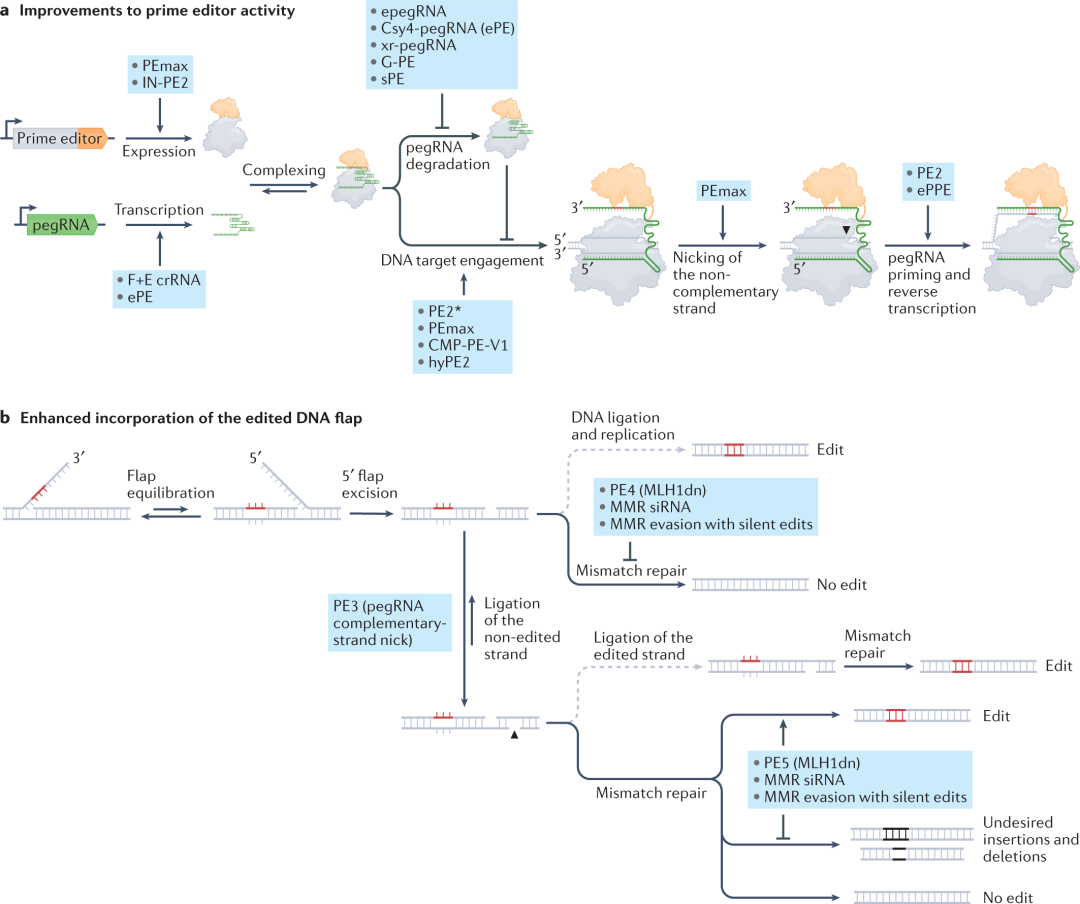
prime editing的成功实现需要许多步骤协调一致。要与靶DNA结合,必须首先表达prime editor,再将其运入细胞核,然后使其与pegRNA形成复合物。接下来prime editor必须有效地找到目标位点,切开暴露的DNA链,并启动pegRNA的逆转录。然后,必须通过细胞DNA修复过程将编辑后的DNA链复制到未编辑的链上,使编辑永久化。因此,提高素prime editing性能的重点是提高这些步骤的效率。
1. 需要增强的prime editors效应蛋白
许多小组通过在Cas9和RT结构域之间插入DNA或RNA相互作用蛋白,优化了prime editor的结构。K. Kim,H. Kim和他们各自的同事在linker内用额外的高迁移率基团肽(CMP-PE-V1)或hRad51 DNA结合结构域(hyPE2)改造了PE,在哺乳动物中比PE2有一定改进。Gao及其同事通过从MMLV RT中移除RNaseH结构域并在Cas9和RT结构域之间插入病毒核衣壳蛋白,类似地构建了一种工程植物原编辑(ePPE),这两种结构域共同将水稻原生质体中的编辑平均增加了5.8倍。未来将致力于表征PE–pegRNA–DNA靶复合物结构从而设计更活跃的prime editor结构,或针对感兴趣的细胞类型或靶点进行优化的定制prime editor变体。
2. 改进用于prime editing的pegRNAs
3ʹ延伸的降解可能导致产生有缺陷的PE–pegRNA核糖核蛋白(RNPs),其与正常的PE–pegRNA竞争结合靶DNA,从而削弱prime editing效率。Liu及其同事通过将RNA结构基序evopreQ1或mpknot添加到pegRNA的3ʹ端,以保护3'延伸不受核酸外切酶的影响。这些工程化的pegRNA(epegRNAs)与经典的pegRNAs相比,编辑效率提高了1.5至4倍。其他小组则在pegRNA 3ʹ延伸中添加了寨卡病毒外核糖核酸酶抗性RNA基序(xr pegRNA)、G-四链体(G-PE)或茎环适体(sPE)。提高pegRNA转录或稳定性也可以提高prime editing的效率。用RNA聚合酶II表达pegRNA也消除了避免poly(U)终止信号的需要。此外,分离pegRNA内的spacer和模板序列也可以改进prime editing。Sontheimer及其同事设计了一个分裂的pegRNA系统,该系统由靶向PE的正常sgRNA和包含primer结合位点(PBS)和RT模板的线性或圆形引物编辑模板RNA(petRNA)组成。
3. 减少prime editing的副产物,改进DNA修复
PE2和PE3b编辑系统不会同时在靶位点切割两条DNA链,因此很少产生indel副产物。Liu及其同事发现,DNA错配修复(MMR)途径强烈拮抗prime editing并刺激indel副产物产生。Loizou及其同事还报告,敲除MMR可提高prime editing效率和精确度。这些启发了两种改进prime editing的方法。第一,设计了MMR蛋白的显性阴性变体MLH1dn,以瞬时抑制MMR并提高prime editing的效率。第二,在预期编辑附近编程额外的沉默突变,可以创建具有更多失配的异源双链中间体,从而避免MMR识别,提高prime editing的效率,但不会全局干扰MMR活动。
4. 染色质可及性能影响prime editing
成功的prime editing需要基因组靶点足够容易被PE–pegRNA复合物接触。Kim及其同事表明,具有染色质调节肽(CMP-PE-V1)的PE变体可以打开靶位点的局部染色质,并改善哺乳动物细胞中的prime editing。他们还发现,通过截短、失效的sgRNA靶向PE以结合编辑位点附近,也增加了染色质可及性和编辑效率。
5. 改造prime editors变体
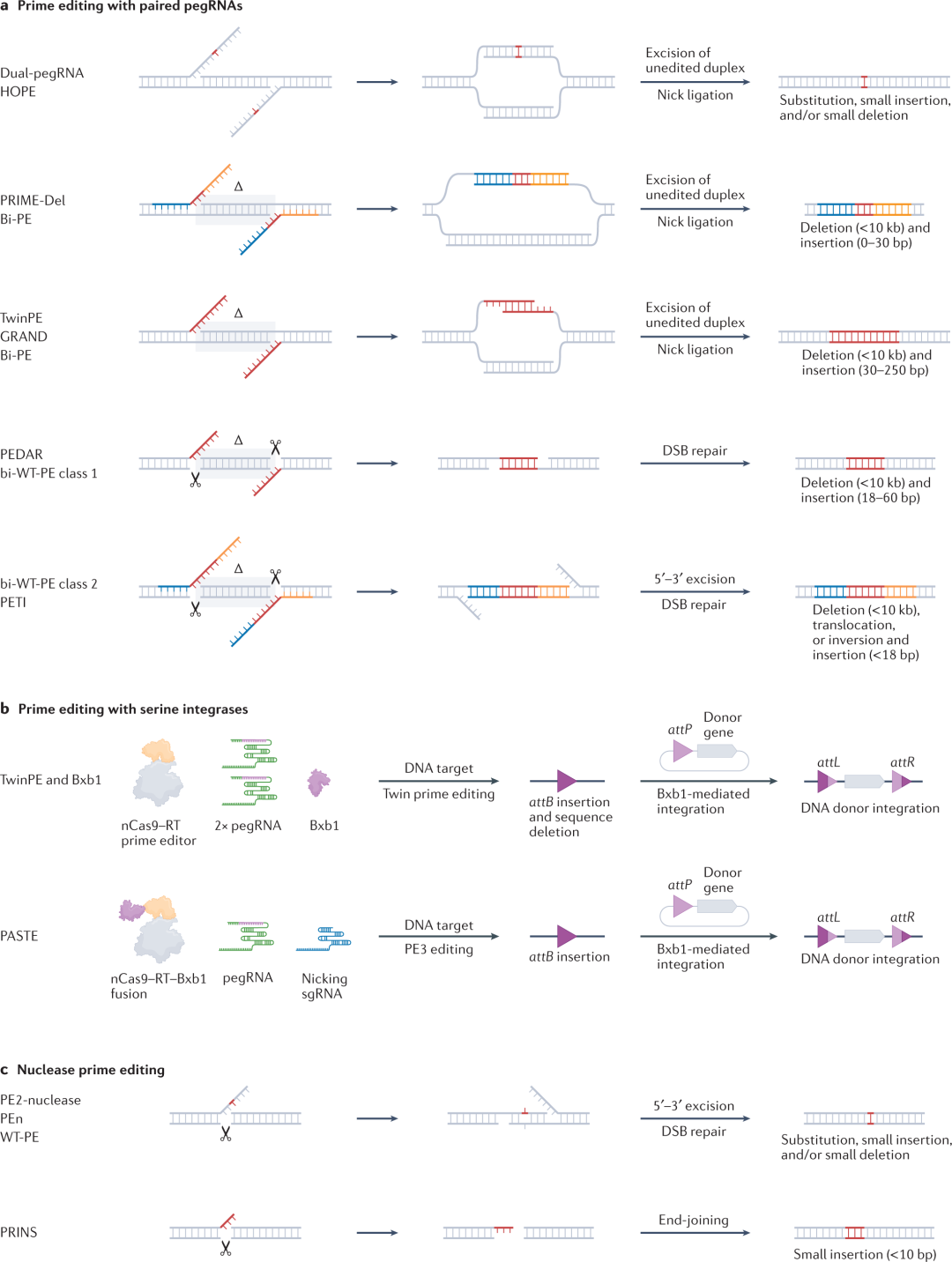
尽管prime editing不需要在靶点精确定位PAM,但编辑远离PE靶点的核苷酸(>40 nt away)仍是挑战。对远距离碱基的prime editing需要创建一个长的pegRNA,合成一个包含编辑的长3ʹDNA flap,并将该flap并入基因组。因此,可能很难编辑远离典型PE酶的NGG-PAM序列偏好的DNA。为了扩大可以有效prime editing的序列范围,研究人员构建了利用具有非NGG PAMs或替代核酸酶切割位点的其他Cas9变体的PE。尽管PE变体扩大了可编辑靶DNA的范围,但仍需要进一步的工作来提高其活性。提高Cas结构域与PE的兼容性十分重要。除了交换Cas9结构域之外,还建立了利用替代RT的PE变体。更小的RT变体也可能产生更小的PE,更容易递送。
6. 预测pegRNAs设计和效率
pegRNAs的优化对于最大化prime editing至关重要,多个小组开发了设计pegRNA的程序。然而,这些工具中只有一些能够准确预测给定靶位点和编辑的pegRNA 的PBS和RT模板长度。Gao及其同事观察到,PBS和基因组DNA链之间的最佳熔化温度在植物中为30℃ ,但本指南可能不适用于37℃培养的哺乳动物细胞 。为此,Kim及其同事在人类细胞中进行了pooled pegRNA–靶位点库筛选,以训练深度学习模型DeepPE,该模型预测最佳pegRNA PBS和RT模板长度。Cheng及其同事还使用Kim等的数据集来推算自己的深度学习模型Easy-Prime。这些计算工具以及未来可能开发的其他prime edit模型,有可能简化prime editing的使用并扩大其适用性。
三、prime editing成分的递送
1. DNA转染和病毒递送
在靶细胞中高效递送或表达PE蛋白和引导RNA是生产性prime editing所必需的。一种常见的策略是将DNA编码的prime editing组分递送到靶细胞中,并利用细胞的内源性转录和翻译机制来产生PE RNPs。在许多培养的哺乳动物细胞系中,脂质介导的瞬时转染或电穿孔prime editor质粒可以介导高效率的编辑,选择转染的细胞可以进一步提高编辑性能。但是,DNA递送具有外源DNA整合到基因组中的风险。
病毒提供了许多递送优势,包括有效的转导和组织选择性,使其成为体内递送PE的一种有吸引力的方式。慢病毒能基因组整合PE,通过插入突变诱导致癌,并通过持续表达增加脱靶编辑。瞬时表达降低了这些风险,因此有利于治疗应用。在整合风险较低的DNA病毒中,腺相关病毒(AAV)、腺病毒和单纯疱疹病毒(HSV)先前已成功用于短暂转导。
2. mRNA递送
与DNA转染和病毒转导相比,PE编码mRNA的递送消除了DNA重组到基因组中的可能性,并可以通过缩短PE表达的持续时间来减少脱靶编辑。PE mRNA和化学修饰或体外转录的guide RNA的共电穿孔,已被证明能在培养的细胞系、原代人T细胞和人干细胞中介导有效的prime editing,在大多数情况下效率高于质粒转染。此外,增强的PE4和PE5编辑系统特别适合通过mRNA递送,这可以限制MMR抑制剂MLH1dn的潜在诱变后果。多个小组还证明,微量注射PE mRNA可以刺激小鼠胚胎中的prime editing,但在没有MMR抑制的情况下使用PE3时可能会产生高水平的indel副产物。因此,PE的RNA递送是体外编辑细胞、胚胎和造血细胞的有效策略。脂质纳米粒子(LNP)也经常用于封装编码基因组编辑剂的mRNA,并将其传递到活体小鼠和人体中,但使用它们传递prime editors尚未有报道。
3. RNP递送
PE–pegRNA RNP的递送可以缩短靶细胞暴露于PE试剂的持续时间,最大限度地减少脱靶编辑。此外,靶细胞中的内源性转录和翻译机制在由RNP递送时不需要产生PE。在第一次用RNP进行prime editing的演示中,Yeh及其同事纯化了与pegRNA复合的PE2蛋白,并将这些RNP微注射到斑马鱼胚胎中,产生了一定的编辑。这些PE-RNP也被电穿孔到培养的细胞系和原代人T细胞中,但所得的prime editing效率大大低于先前从质粒或mRNA递送中观察到的。其他人也观察到永生细胞系和人iPSCs中PE RNP的编辑效率同样低。因此,尽管RNP递送提供了理论上的优势,与DNA和mRNA递送相比,当前的PE-RNP递送方法在效率上滞后。
四、prime editing的应用
1. 研究并治疗疾病
生物医学界对使用prime editing来建模并纠正导致人类疾病的基因突变表现出浓厚的兴趣。约90%的人类致病性遗传变异是不到十几个碱基对的单碱基突变或插入和缺失,prime editing系统能够处理这类DNA变化类型。prime editing还可以进行精确的基因操作,间接改善疾病或为疾病提供保护。许多人类疾病如冠状动脉疾病,都是多基因疾病。大多数基因组编辑工具缺乏灵活性,无法同时编程多个精确的DNA变化,但prime editing可以使用pegRNA的串联阵列进行几次所需的编辑。此外,相同的PE治疗有可能治愈具有不同突变的异质患者群体。早期的体内实验表明,prime editing有可能纠正动物体内的致病性突变。尽管prime editing在纠正人类患者遗传疾病方面具有很大潜力,但将其输送到其他器官,如心脏、肌肉、肺和中枢神经系统,仍然是治疗性prime editing应用的主要挑战。创新递送技术并结合上述PE蛋白和pegRNA组分的最新改进,对于最大限度地提高动物的prime editing效率至关重要。
2. 生成动物模型
对模式生物的DNA进行工程设计,有助于阐明基因及其编码的生物分子在复杂系统中的功能。为了生成具有所需基因型的动物模型,基因组编辑通常在早期胚胎或生殖细胞中进行,以最大限度地减少嵌合的机会。prime editing比使用CRISPR–Cas核酸酶的HDR显示出更高的编辑纯度,并且比碱基编辑具有更大的编辑灵活性,使其非常适合创建具有精确基因组变化的转基因生物。prime editing可以促进高精度转基因动物模型的创建,有效的胚胎prime editing有助于对动物的多基因特征进行多重编辑,并允许研究人员直接对动物进行表型编辑,而无需异型杂交(outcrossing)。
3. 筛选遗传变异的功能
引入遗传扰动(genetic perturbations),并通过功能表征或富集评估其后果是揭示生物途径和机制的普遍方法。由于精确性和多功能性,prime editing非常适合用于功能性筛选的基因突变。通过对编辑后的基因座进行测序来鉴定prime editing所产生的DNA变化,将突变区域限制在单个基因上。相反,对pegRNA进行测序也可以恢复编程的prime editing的身份,但不会对编辑的位置施加限制。prime editing系统的不断进步将有助于跨多组分途径对基因变异或甚至基因变异组合进行遗传筛选。
4. 标记内源性基因
标记内源性基因的能力广泛用于选择性地检测或操纵细胞中的蛋白质。原始PE3 prime editing系统可以编程插入18 bp的6 × His标签或24 bp的FLAG标签(DYKDDDDK),还可以安装类似大小的其他标签,如HiBiT荧光素酶、GFP或细胞定位信号。此外,新一代prime editing系统已被证明可以介导更大标签的安装。这些结果建立了将蛋白标签或肽段安装到感兴趣的内源性基因上的prime editing能力,提高PE的长片段插入效率将解锁更多的基因标记应用。
结论和未来展望
以可编程的方式精确操纵基因组,改变了生命科学研究。与其他基因编辑技术相比,prime editing提供了更大的通用性、产物纯度和靶序列特异性。prime editing的潜力推动了许多创新研究来提高其效率并扩大其能力,展示其在基础研究和治疗中的应用和无限可能。
原文链接[已授权转载]:
免责声明
本文旨在行业资讯和知识分享,无任何商业用途。文章仅代表作者观点,不代表本站立场。

*CUT&Tag实验笔记来啦~小翌给你准备了
