
UT&Tag(Cleavage Under Targets and Tagmentation)是蛋白-DNA互作的一大革新技术,它不需要使用甲醛交联以及免疫共沉淀,而是通过针对靶蛋白(如转录因子、染色质重塑蛋白)的抗体和Protein A/G的介导,使得与Protein A/G融合的Tn5酶(Tagmentase)在切割DNA片段的同时在序列两端加上测序接头,经PCR扩增后即可形成用于高通量测序的文库(见图1)。CUT &Tag技术具有细胞投入量低、信噪比高、实验周期短(1天实现从细胞到文库构建)、可重复性好等显著优势,尤其适用于早期胚胎发育、干细胞、肿瘤以及表观遗传学等研究领域。
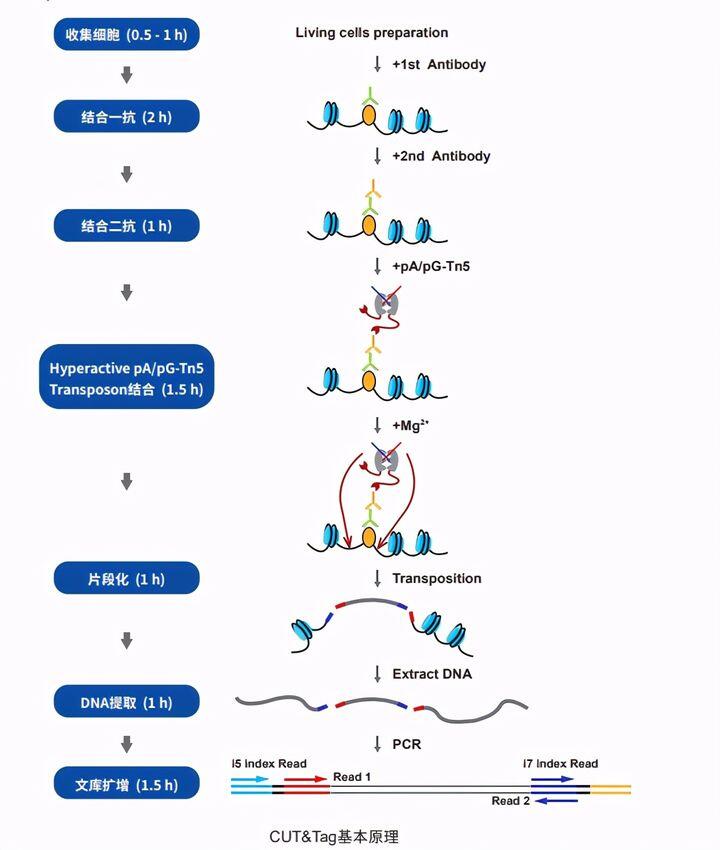
由于CUT&Tag主要是针对极低细胞起始量进行实验,因此对核心酶原料有着极高的要求。CUT&Tag技术的核心原料酶是Hyperactive PA/PG-Tn5 Transposase,它具有高活性,对微量DNA有高灵敏度和高亲和力,能有效抓取数十个细胞中的有限结合位点等特点。
Hyperactive PA/PG-Tn5 Transposase结构
Hyperactive PA/PG-Tn5 Transposase的N端结构域为金黄色葡萄球菌A蛋白(Protein A) 的一部分或链球菌G蛋白(Protein G)一部分,C端结构域为Tn5。两个结构域通过短的刚性连接肽(Linker Peptide)连接,使得融合蛋白酶兼具PA/PG &Tn5活性。通过针对靶蛋白(如转录因子、染色质重塑蛋白)的抗体和Protein A/G的介导,Tn5 Transposase可以实现切割靶区域的目的。
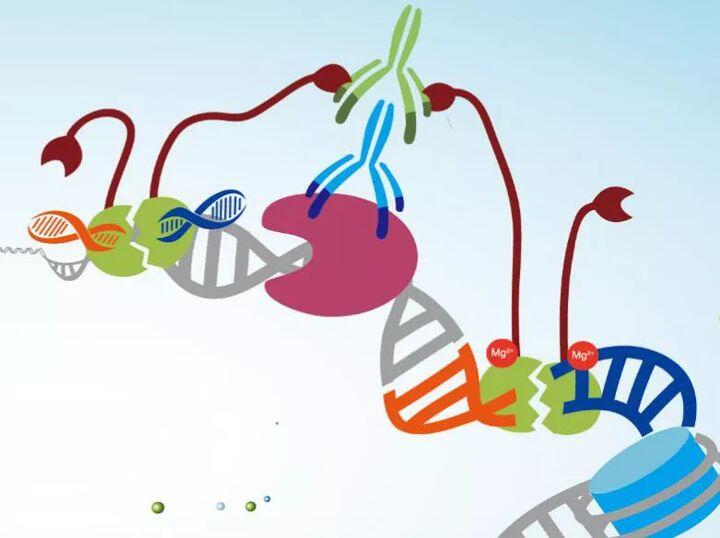
Protein A/G
Protein A 来源于金黄色葡萄球菌的一个株系,它含有5个可以和抗体IgG分析的Fc段特异结合的结构域。Protein G是一种源自链球菌G蛋白的细胞表面蛋白,为三型Fc受体。与Protein A类似,Protein G也可以与IgG的Fc段特异性结合,不同的是,Protein G还可以和某些抗体的Fab和F(ab’)2段结合。
Tn5转座酶
转座酶是一大类细菌来源的蛋白,通过结合转座子序列末端并催化其转移到基因组上随机位置,实现“剪切-粘贴”或“复制-粘贴”型的DNA插入的作用。其中Tn5转座子因其转座的随机性好、稳定性高、插入位点容易测序等特点,已经成为分子遗传学及基因诊断学研究的热门工具。
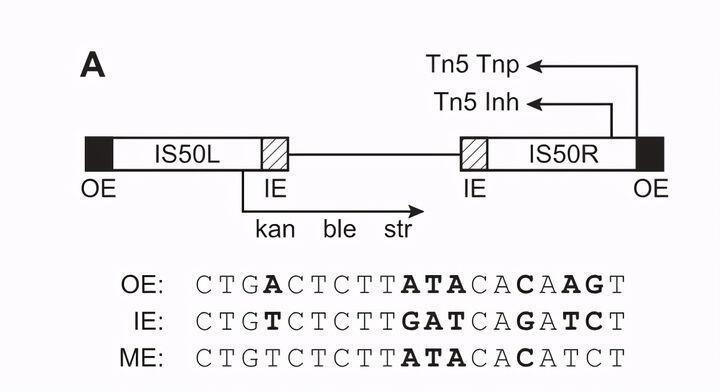
Tn5转座子是一种细菌转座子,全长约5.8 kbp,由编码三个抗生素(新霉素、博莱霉素、链霉素)的核心序列和两条倒置的IS50序列组成。其中IS50R和IS50L的序列高度同源,只是IS50L的一个碱基存在突变。IS50具有19bp的倒置末端(ES),外末端outside end(OE)和内末端inside end(IE),两末端倒置有7个碱基不同。此倒置末端是转座酶(Tnp)的作用位点,在转座过程中,将转座子从供体DNA中完全切除,并插入到目标DNA中。IS50L和IS50R均含有编码转座酶(TnP)以及转座阻遏蛋白(lnh)的基因,但由于IS50L中的碱基突变,造成翻译提前终止,所以仅有IS50R可以产生正常的有活性的TnP和lnh。
60 bp的转座子底物由20 bp的dbb( ES-donor backbone,ES供体主链),19 bp的ES和21 bp的转座子组成。

Hyperactive PA/PG-Tn5 Transposase作用机制
Hyperactive PA/PG-Tn5 Transposase利用PA/PG寻找不同来源的抗体,结合在靶蛋白的附近后再进行切割。它的切割作用由Tn5转座酶实现,Tn5转座酶作用机制主要分为三步:
01:转座酶(Tnp)分子(红色)与转座元件末端与特定的19 bp ES识别序列(白条)结合,形成两个TnP-ES复合体。随后两个TnP-ES复合体通过TnP的C末相互作用进行联会,形成Tn5转座酶复合体。
02:在Mg2+存在下,Tnp活化水分子,活化的水分子进行亲核攻击,水解DNA的一条链,从而在转座子末端暴露出一个OH基团。然后激活此OH,以对DNA的互补链进行亲核攻击,形成发夹结构并切割DBB,并产生DEB复合物(具有两个切割末端的联会复合物)。每种单体都与它自己的ES(顺式接触)和与其他单体结合的转座子ES都进行接触(反式接触)。每个Tnp的催化结构域以这样的方式定位:切割另一个单体的转座子末端(反式切割)。
03:在DEB复合物与非特异性靶DNA结合后,转座子末端的活化的3′-OH基团对靶DNA进行亲核攻击,以9 bp的间隔完成链转移。Tnps离开目标DNA,缺口被修复,产生9 bp的重复。

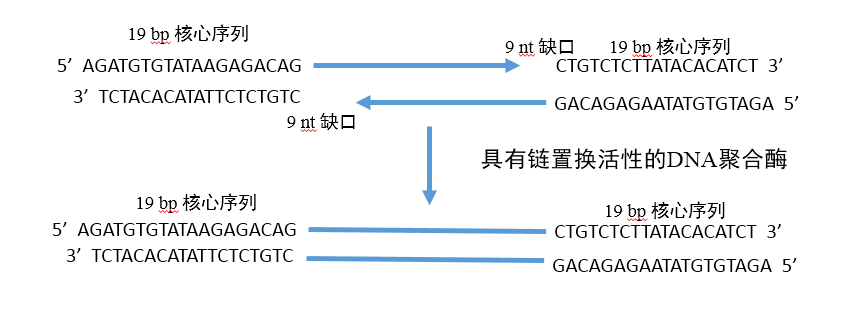
研究人员发现,整个转座子序列并不是转座必须的,只需转座子的19 bp末端核心序列,转座酶便能将该部分序列插入并连接至基因组内。利用这个原理,将测序接头序列加入到末端核心序列中,在转座酶进行片段化的同时,加上了测序序列,省去了接头连接的过程,只需在片段化后进行PCR扩增便可得到完整的用于测序的文库。但是这一步之后,接头与插入片段之间有9 bp的gap需要补平(如图6),这就是为什么Tn5建库在PCR之前必须进行72℃反应3min,而且所用的PCR酶需是具有链置换功能的非热启动酶。
传统建库方式需要经过DNA片段化、末端修复、接头连接、文库扩增等步骤,耗时较长,而使用Tn5转座酶进行文库构建,可将DNA片段化、末端修复、接头连接等多步反应转变为一步简单的酶促反应,极大缩短建库时间,提高工作效率。

Vazyme Hyperactive PA/PG-Tn5 Transposase优势
切割活性高
野生型Tn5转座酶的活性很低,以减少对宿主产生致命突变的风险。Tn5转座酶活性很低的部分原因在于其蛋白的N端和C端在三维空间构象上相互接近,并发挥相互抑制的作用。诺唯赞专注于酶改造,使用基因工程手段对于N端或C端的突变,可以得到高活性形式的突变形转座酶。使用Vazyme pA-Tn5与市面上A公司同类产品使用50 ng gDNA 在55℃条件下打断10 min后,扩增9个循环,利用Qubit检测浓度比较转座酶活性,发现Vazyme PA-Tn5切割活性远远高于A公司同类产品。

核酸残留低
由于Tn5转座酶对DNA有极高的亲和力,导致了纯化过程中易产生非特异核酸残留。Henikoff实验室设计不同投入量细胞进行CUT&Tag实验,并分析不同投入量下E.coli残留情况。发现随着细胞投入量下降,E.coli reads 越多。
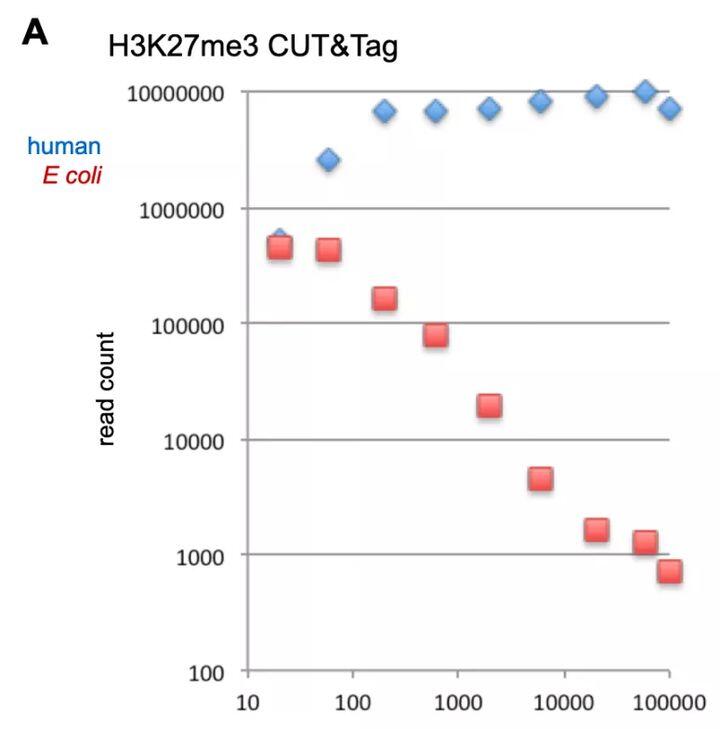
诺唯赞生物针对Tn5酶非特异性核酸残留问题进行攻克(见图11),优化酶生产工艺,大大降低了低起始量细胞建库中非特异核酸占比。
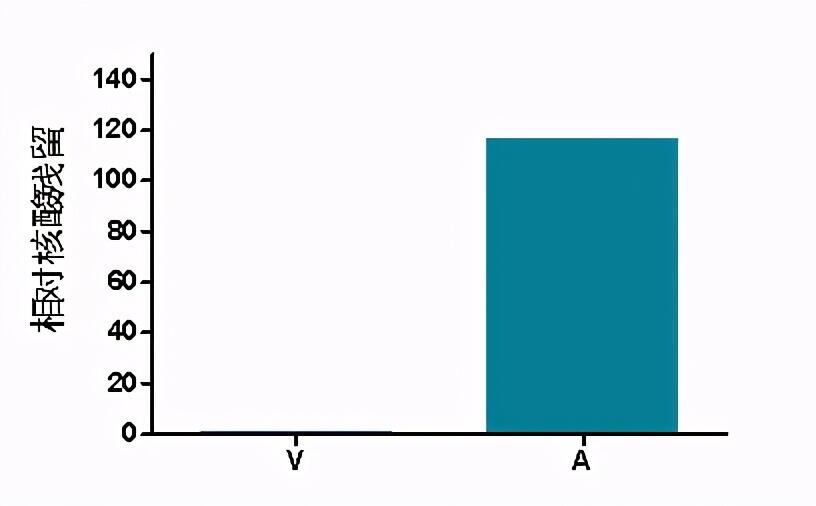
核酸残留检测方案:实验人员使用PA-Tn5裸酶进行PK消化,富集并回收核酸。设计针对细菌23S通用qPCR引物,对回收的核酸进行qPCR检测,计算E.coli核酸残留。
