

进化分类学浅析
作者:bilibili热爱古生物的蔡文姬
分类学科普确实很难,因为它有很多不同的流派,各种派别之间的观点既有联系也有区别,并且现在在实际应用中还在相互渗透,所以相当多的人只知其一不知其二,或者是只记住个结论,然而这个结论怎么来的,依据的标准是什么,就不清楚了,所以今天我就来把上世纪中后期开始比较流行的一个学派,也是现在实际应用较为广泛的一个学派——进化分类学派的一些观点来简单介绍一下。虽然这样的分类学科普很大程度上是枯燥乏味的,但是我相信有感兴趣的朋友会看进去的,因为确实也没法讲的很有意思,概念性的东西很多,加上我这个人讲话比较正经,所以其实不求能有多少人看得进去,只求喜欢的人真的能有收获。
首先,进化分类学属于“谱系分类系统”的一种,以进化论为基础进行分类,即分类结果要符合生物进化的系统发育谱系,反映亲缘关系。但是较早期的分类学除了系统树泛滥之外,进化论并没有起到什么推进作用,该怎么样还是怎么样一切照旧,直到上世纪三四十年代,关于分类学的三大问题逐渐被重视起来,成为了讨论的焦点,当时的情况是百花齐放百家争鸣,分类学也借此环境得到了良性的飞速发展。那么三大问题是什么呢?分别是物种概念,系统原理和特征分析,下面我分开详细讲述一下。
一 物种概念
不要看现在有关生物“种”的概念以及分类学基础在中学课本甚至小学课本也都反复讲过,但是有关物种的概念确实不是一句两句能说得清楚的。早先林奈时代的观点是,物种是不变的,当初上帝创造出多少物种,物种就有多少种,不会出现新种,进化论打破了这一观点,按照进化观点,种与种之间是历史上连续的,不是间断的,因此物种应该是变化的,这是进化论对于旧观点的重大冲击,也是认识上的进步。
但是说来奇怪,虽然林奈的“物种不变”早就被人们抛在了历史里,但是当时他鉴别并确认是种的动物现在很多依然认为是种,尽管种以上的纲目科等单元变化了又变化,到现在基本和当时已经面目全非了,但是种一级却相对变化很小。此外我们现在在对物种的鉴别,一是要求特征分明,即有足够的独立特征区分于其他种,二是要求特征不变,变化的特征不能够确定种,这样一来,物种之间的界限不就很清晰了么,物种又成了不变的。进化论指出了物种是不断变化的,但是这种变化本身否定了物种的存在,分类学中肯定了物种的存在,但是依据却是物种不变,这样就产生了变与不变的矛盾,于是人们就在想,物种到底什么?它是否真实存在?还是人们为了分类方便而设立的分类单元呢?如果它存在,生命的一代一代是连续的,但是不同物种之间的区分又是严格的,那么它到底是连续的还是间断的?
因此有关物种到底存不存在的争论曾经持续了很久,直到现在有关于“什么是种”的概念也难以统一,在不同的学派中“种”有不同的定义,现在只给出分类学家的结论吧,进化分类学中的观点认为,“种”是又变又不变,又连续又间断的。关于具体的“种”的不同概念,以及各有怎样的意义,我们放到以后的专栏里结合其他内容一起谈一谈,因为这个问题是任何分类学,或者只要谈生物进化就绕不开的共性问题,现在谈的话概念性的东西就比较多了,而且对于了解进化分类学来讲,主要还是明白其操作的系统原理以及如何分类,在此仅简单点明一下。
二 系统原理
进化分类学要求,分类结果应该反映正确的系统发育关系,强调物种之间的亲缘关系和演化关系,而非按照表型机械归类,下面就来谈一谈分类学依据的系统原理。
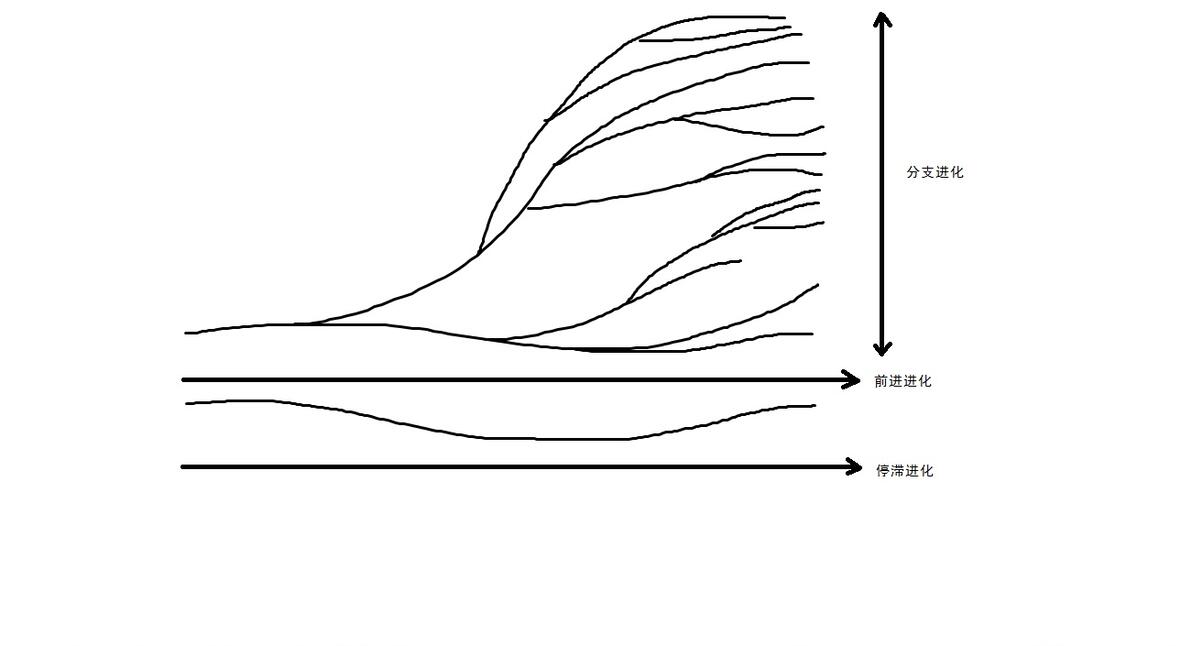
首先,一个类群的出现,是从无到有的变化,大到植物界动物界,然后小一点的,哺乳纲鸟纲,再小一点的真兽下纲,再小的猫型亚目等,它们的起源发展都是这样一个从“无”这个类型的生物到“有”这个类型的生物的变化,这一整个类型的生物有一个共同祖先,由这个祖先演化出形形色色的其他生物。
而一个类群无论其大小,在进化上都有着横向和纵向的两个发展方向,横向的分支发展产生物种的多样性,是由少到多的发展,纵向的阶段发展产生结构的复杂性,是从低到高的发展。进化分类学的系统原理便是对这几条理论的概括,从无到有,从少到多,从低到高,可以概括为共同起源,分支发展,阶段发展。
相关类群定义
其实这几个概念从字面上就很好理解,并且相互之间是有联系的,也不好分太开说。先说共同起源,顾名思义共同起源就是要求一个类群要有一个共同祖先,即这个系统是单源系统,分类学要求建立单源系统,这样才算自然类群,而拒绝多源系统(多种起源,多个祖先)。鉴别单源系统,对于现代生物来讲,可以经过特征分析(后面会说),或者现在有了分子生物学技术,可以非常方便地确定它们之间的亲缘关系从而建立出较为可信的系统发育谱系。而古生物只有化石依据,理想的结果是,找到过渡类型的化石,便可以很方便确定一个类群的祖先是谁(不一定是具体的哪一种,也可能是哪一类),比如,我们现在都知道,鸟类是从恐龙起源的,但是在上世纪初期的时候,因为化石证据不足,这一点并不明确,人们虽然肯定了鸟类和爬行类之间存在亲缘关系,但是并不能正确认识到鸟类到底是从爬行类的哪一个阶段,或者哪一个类群里起源的,所以才有了曾经盛极一时的直到现在也有少数学者在坚持的槽齿类演化说,认为鸟类和鳄类,恐龙,翼龙一样起源于古槽齿类(用支序分类学的术语来讲,基位主龙形类)。这种学说当时和恐龙起源说谁也说服不了谁,直到后来始祖鸟的发现,以及越来越多的羽毛恐龙的发现,强烈地显示了鸟类和手盗龙类恐龙的亲缘关系,这样我们就知道了鸟类是来源于恐龙的。如果没有确切的过渡类型或祖先类型化石,那么对于一个类群便要依靠特征分析来进行甄别,需要指出的是,我们说的是因为它们具有亲缘关系所以具有相似特征,而又说因为相似特征把它们归成一类,这样是不是又出现了转圈话的矛盾呢?其实不是的,特征分析也是有单独的原理的,并不是任意一个相似特征都可以拿来当做分类依据,比如没人会因为都是恒温动物而把鸟类和哺乳类划分为一个分类单元(注意是进化分类学上的“分类单元”),特征分析的具体内容会在稍后的篇幅描述。
然后是分支发展,分支发展产生物种的多样性,一个新类型的出现,理论上是由一个物种,经由分化,由少变多,产生多种多样的类型,形成一个或大或小的自然单元。这个其实举例子的话,加拉帕戈斯地雀就是非常好的例子,正是加拉帕戈斯岛上的13种地雀,启发了达尔文思考物种进化。这个例子应该在课本上有,但是我还是简单复述一下,这13种地雀,大小相似,但是生活习性各不相同,有地栖的有树栖的,有食虫的有食种子的等,区分它们的关键是是喙的形状各不相同,人们相信它们是由一个祖先演化来的,后来经过一系列的环境适应产生了不同的喙型(实际上,是先有了喙型变异再经历适应环境的自然选择,而不是为了适应环境去进化出不同的喙型),并且后来的研究也证实了这一点,这13种地雀加上科科岛上的另一种,一共14种地雀有一个共同的祖先种,经过地理隔离,生态隔壁,逐渐分化产生了不同的类型,它们共同组成了一个自然类群,这就是一个分支发展的过程。
往简单了说,分支发展就是在一个共同祖先的基础上,经过不同的隔离分化,从一个种分化为能适应不同的空间,产生不同的生活习性,占据不同生态位的多种生物的过程,这个祖先和它产生的所有分化出现的种类共同组成一个自然分类单元,显然地,这种分类单元是单源的。这种一般叫做“辐射适应”,这个词我的理解是,从一个物种分化出多个适应不同环境的物种,就像辐射一样,宽度突然就铺开了,应该是十分形象的。比如石炭纪的两栖类的辐射适应,中生代的爬行类辐射适应,新生代的哺乳类辐射适应等。无论类群大小,无论演化程度高低,任何一个单源自然类群都是从一个祖先(不管现在是否能找得到),经过了一次或多次的辐射适应而产生的。这种分支进化的特点是,在分化初期不产生机体的复杂性变化,而是着重于对不同环境的适应,产生形状的分化,进而产生新的种类,其基本过程是“种群分化-物种分化-类群分化”,也就是说分支进化是产生物种多样性的基础。
最后说阶段发展,有了上面分支发展的基础,阶段发展也就好理解了,分支进化在横向上产生物种的多样性,而阶段发展在纵向上产生物种结构的复杂性。从大的方面,生物的发展经历了几次大的阶段,从无细胞结构到有细胞结构,从原核生物到真核生物,从单细胞生物到多细胞生物等,可以看出来,生物的进化是向着结构功能越来越完善和复杂的方向进化的,仅仅靠分支发展无法体现这一点,所以在分类学中,除了分支发展,还要有阶段发展的考量。简单说就是,在分支内,还要考虑不同分支的物种的进化程度,以主龙类来讲,主龙类的两条主要支系鳄鱼和鸟类(不考虑恐龙,只考虑现存),其中鸟类相比于鳄鱼,显然是进化速度较快的,其相对于祖征演化出了更多的衍征,而鳄鱼则相对保留了较多祖征,进化分类学家们认为,考虑到阶段发展,鸟类的进化程度明显要高于其他的爬行类,因此可以考虑作为新的类群起点从爬行类中分离出来,以体现不同类群的进化程度不同,只考虑分支发展的话,这两个类群被认为是平行的,无法体现结构的复杂化。不过这一点支序分类学派的分类学家认为是不正确的,这个我们留到后面对比的时候再说。
这样我们就说完了进化分类学的三条基本系统原理,其实这三条是相辅相成的,比如我们构建这样一组系统发育图,1-4代表四个不同起源,其中每个起源都是分支分化的开始,同时也是阶段复化的开始,起源产生分支,分支产生阶段,阶段是新的起源,这样就把这三条系统根原理统一了起来。
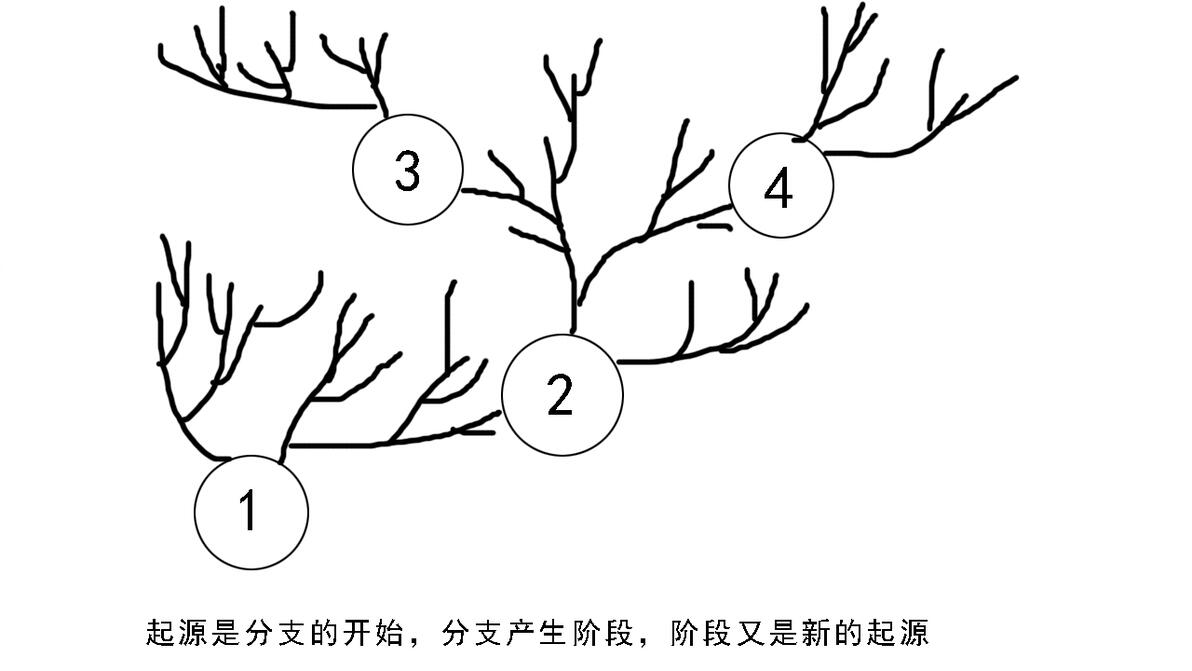
以四足动物进化为蓝本的系统发育简图
三 特征分析
其实主要想讲的部分是系统原理,搞清楚了原理才能用原理去指导分类实践,对于进化分类学的特征分析,实际上也有很多需要考量的点,在此主要详细讲述常用部分——共性和特性,祖征和衍征,这两种也经常被用在支序分类学中来区分不同的分类单元,其他如个体群体等不做详解,因为一谈多了又要涉及到前面物种原理的变与不变,间断与连续之间的关系了,这些辩证关系部分我不想在这里多提,主要还是讲一下用得多的实用部分。
特征分析,顾名思义就是对物种的特征进行辨别分析以便于将具有相同或相似特征的物种归类,其中最基本的原则是,对立特征才有分类学上鉴定物种和归类的意义,非对立特征是无意义的,分类决定于构造的对比,不决定于构造本身。也就是说,至少要有一对互相对立的特征才能成为鉴别分类的基础,比如,鱼类相比于四足类,就有具有带有脚趾的四肢和不具有带脚趾四肢这样一对对立的特征,以便将这两个类群区分开来,如果只考虑鱼类本身,所有的鱼都不具有脚趾,或者考虑所有的四足动物,没有不具有这种特征的(蛇算退化了),这种就没有分类意义,因为要不就全有要不就全没有,没有分辨价值。
实际上共性特性,祖征衍征可以看成是类似的东西,或者说,祖征就是共性,衍征就是特征。共性和特性其实为分类单元的建立提供了有力的鉴别标准,经过这样的层层分类,每一个阶段被分的清清楚楚。还是以上面的例子为例,简单谈一下,我们将四足动物与鱼类分开的依据是,前者具有脚趾,可以上岸生活, 可以使用肺呼吸等,这些特征和鱼类对应的特征是对立的,所以将其与鱼类分别了出来,对于整个脊椎动物来讲,上述特征相对于基础类群鱼类,就是衍征,而在考虑四足动物内部的分类时,这些就成了共性,成了祖征,便不再具备继续分类的特征,而要考虑其它的更加衍化的特性,又如在这群四足动物中,有一部分动物可以完全离开水,完全使用肺呼吸,并且演化出了羊膜卵结构,这些结构和特征是两栖类不具有的,因此这些又作为衍征,成为区分爬行动物(羊膜动物)与两栖动物的依据,后面的鸟类哺乳类就不展开了,都是一样的过程。概括一下就是,上一级的衍征是下一级的祖征,上一级的特性是下一级的共性,做特征分析时依据是对立的特性,衍征(或者说是否具有同源的共有衍征)来建立单源系统,而不是祖征(共性),要以能够与其他类群区分出来的二元特征来建立相互排斥的分类单元。系统发育的建立,主要解决两个问题,一从谁那来的,二怎么来的,祖征能够帮我们解决第一个问题,如始祖鸟的祖征,让我们知道了鸟类是从恐龙那里来的,至于怎么来的,就要分析各类过渡类群的共有衍征了。
对于上下单元来讲,衍征的出现体现了阶段发展,祖征的保留体现了分支的延续,是上下级的套筒性质,对于平行单元来讲,衍征的区分则表现了类群之间的对立以及相对祖征的缺失,是平行的对立性质,这与我们之前讲的阶段发展和分支发展的特色也是一致的。鉴别祖征和衍征确立分类单元其实是个很复杂的工作,不能只鉴别几个特征就完事,需要大数据的比对,所以现在可以理解为什么虽然鸟类和哺乳类都是恒温动物但是没人把他们归类在一起了吧,因为其他的关键特征区别太大,恒温既不是祖征也不是它们的同源衍征,是分别独立演化出来的,因此不具备成为一个分类单元的条件。
总之进化分类学的一些主要内容就是这样,当然有很多我没提到的东西,只是挑了一些我个人觉得比较重要又好理解得到部分简单谈一下,后面我来说一下和支序分类学的区别以及二者的优劣。
首先,这两者都是“谱系分类”,都要求建立符合系统发育的分类体系,这一点是一样的,但是他们对于“单源”的理解不一样,对于进化分类学来讲,单源意味着一个分类单元要有一个共同祖先,不接受多源系统,而对于支序分类来讲,他们不光不接受多源系统,而且也不接受同源系统,即单源必须是严格单源,所有后代都要包含进去,进化分类学是不要求这点的,这是一个重大区别;由此引发的下一个区别就是,支序分类学只看共同起源和分支发展,不看阶段复化,因此分支图只能看出系统关系,看不出阶段复化,一个类群的生物,不管你再怎么演化,是变得更高级了也好更复杂了也好甚至面目全非了(比如从始祖合弓兽的形态到智人形态),你也要包含在这个单元里,和其他生物一视同仁,而进化分类学不这么想,进化分类学里虽然也有“阶段”和“分支”的概念(这个概念后来被支序分类学吸收了),但是却认为应该把明显结构复杂化的单独作为新的起点,把演化程度较高,较为复杂的生物还包含在原始类群里是不能被接受的,因此他们接受同源并系群(不是所有的并系群都能被接受,必须是具有共有衍征的单源系统),如传统的很多作为过渡的分类单元都予以保留,这些单元或许不是一个严格的“分支”,但是却是一个从简单到复杂的“阶段”,后来这个概念在支序分类学里也经常被提到,可以体现出二者之间的相互融合。
至于各自的优缺点,自然优点是共通的,就是区别于旧时代的“群魔乱舞”,每个人的人为分类标准不同导致的分类混乱,他们的分类系统都反映了物种本来的亲缘关系,建立的都是各自学说下的“自然类群”,是客观的。不同的是支序分类学不考虑人为判断的复化标准,一切只看分支,而进化分类学涉及到人为判断简单复杂,这一点上就不够客观,可能有不同的人看法不同,比如在恐龙和鸟的交界处,什么算恐龙什么算鸟呢?没有具体标准。但是实际上,支序分类学在建立分支图时也需要大量使用特征分析,既然涉及到特征分析就离不开人,相当多的类群定义(包括节点的选择,特征的选取)还是人定的,虽然它成功解决了什么算恐龙的问题,所有的鸟类都是恐龙,没有争议,但是没有解决什么是鸟的问题,还是要有一个节点来指示“鸟”这个单元的存在。
所以现在很多人建议全部使用全/冠群来定义类群,所谓的冠群也就是包含一个类群现存物种的最近共同祖先和它的所有后代,这样就没有人为标准了,一切看自然谱系,用这样定义的哺乳类就是“人和鸭嘴兽的最近共同祖先和它的所有后代”,但是这样定义虽然是解决了选点的争议问题,但是有两个问题,一是这样定义将范围大幅度收缩,尤其是人们熟悉的鸟类,收缩到冠群之后,原来的反鸟将不再是“鸟”,鱼鸟黄昏鸟也都不再是“鸟”,始祖鸟就更别说了,它们就都成了“非鸟恐龙”,非常不利于人们交流和科学研究,第二个问题是,冠群定义缺乏特征分析上的意义,因为它完全依靠亲缘关系来定义,并且以现存物种为基础,那么就出现了,干群羊膜类不能叫羊膜动物,即使它有羊膜,干群四足类不能叫四足动物,即使它长了腿等等这种看上去比较乌龙的现象,这个毕竟不同于能哺乳的蜘蛛这种跨了纲的,冠群四足类的上一个节点分支如果还活着,那么它按照冠群定义他也是四足类,就因为它死了所以就开除籍贯了?是不是有点别扭。还有就是如果有一天鸭嘴兽没了,假设过几百年之后鸭嘴兽没了,那么这个哺乳类冠群将收缩到后兽和真兽类,鸭嘴兽到兽形类之间的一大票支系包括鸭嘴兽本体突然开除哺乳籍,所以说其实还是有问题,因此,现在很多地方都是两套体系并存,哪个方便用哪个,以免造成不便,不过在描述科学分类时候还是以支序分类为主,毕竟是当下的主流学派。
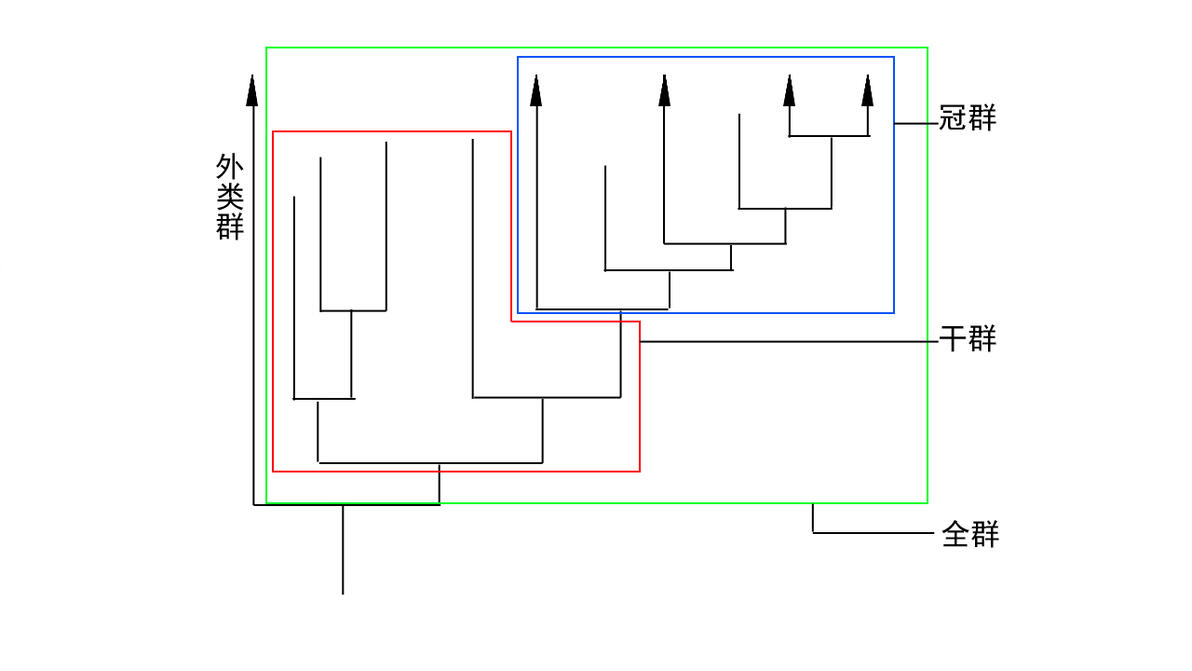
什么是冠群
其实笔者认为,进化分类学虽然在选点的时候会有人为因素,难以统一,但是它有一个绝妙的好处,这也正是支序分类不具备的,就是它的阶段发展原理让我们很清晰认识到了地球生命发展的历程,鱼类-两栖类-爬行类-鸟类和哺乳类的进化链条强烈地反映了脊椎动物各个类群之间的联系和区别(虽然这个链条在支序分类学里认为是不正确的,从所谓的“鱼类”进化到人,并不经历两栖类和爬行类阶段),清晰明快地反映了类群的演化趋势,以及各个类群之间的结构对比,各个类群的进化程度,相比祖先的复化程度在进化分类学的系统关系上一目了然,有助于人们对进化历程有更深刻的认识和总结,所以在平时交流场合,我更建议大家不必拘束于所谓的严格词汇,怎么能够解释清楚怎么来。眼下虽然支序分类学依然是绝对的主流地位,但是其已经在接受其他学派的主张了,包括“阶段”和“分支”的区分和使用,数值分类学派的特征分析手法,并对一部分并系群进行了命名等,正如分类学的目的所言,分类学是一门实用科学,既要反映历史又要方便实践,我们根据场合适当做一下调和,也没有什么不好。
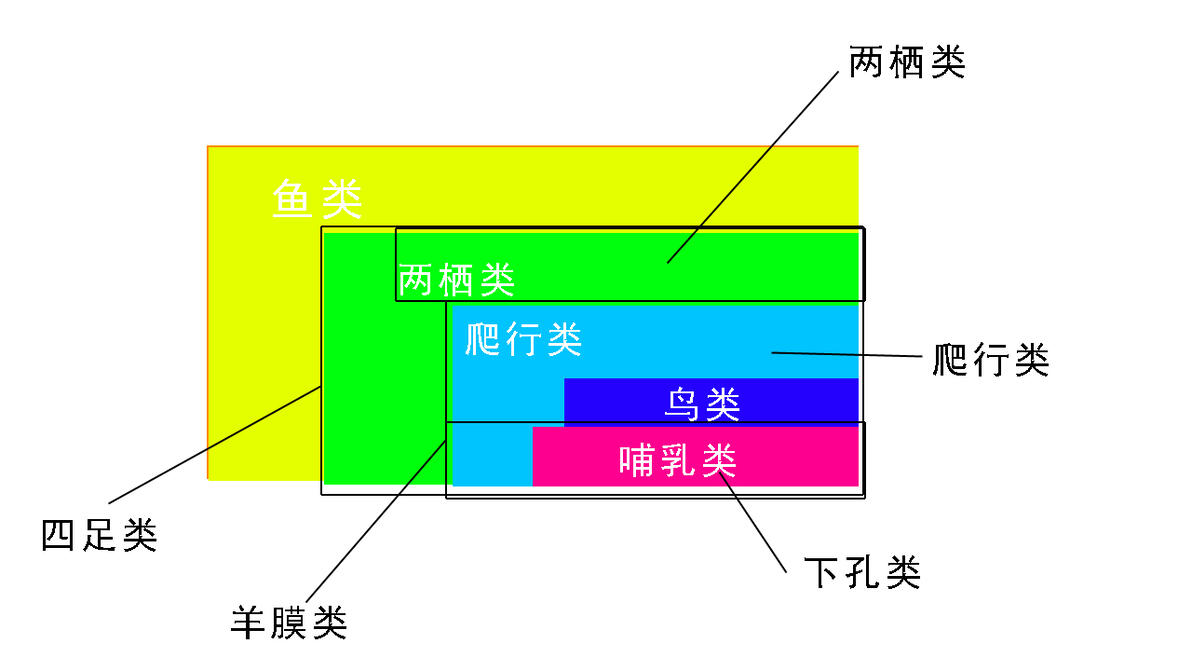
相关类群不同系统的区分
以上就是我对于进化分类学的一个总介入门,资料多采用陈世骧先生的《进化论与分类学》,后加少许自身思考,希望大家能够有所收获,有什么问题可以留言,我能解答的话会回复。
本文仅发布于bilibili,禁止未经许可转载或摘编,如要转载请联系作者并注明出处。
