
本章介绍的是光合作用的后半部分,碳反应。
长期以来人们认为基质中的这些反应不依赖与光,所以称之为暗反应(dark reaction),但是基质中的反应依赖于光反应的产物ATP与NADPH,并且相关的酶活性需要光调控,所以现在改名字了——碳反应(carbon reactions of photosynthesis)。
本章节up将细说光合作用碳反应的三个部分:
由羧化酶催化的光呼吸改变光合作用效率的机制;
植物富集二氧化碳的生物化学机制;
淀粉与蔗糖的合成。(此部分不做介绍)
其中第二条,富集CO2,还存在着与降低植物光呼吸负面影响相关的C4代谢(C4 metabolism)和景天(科)酸代谢(crassulacean acid metabolism,CAM)两个“异端”。
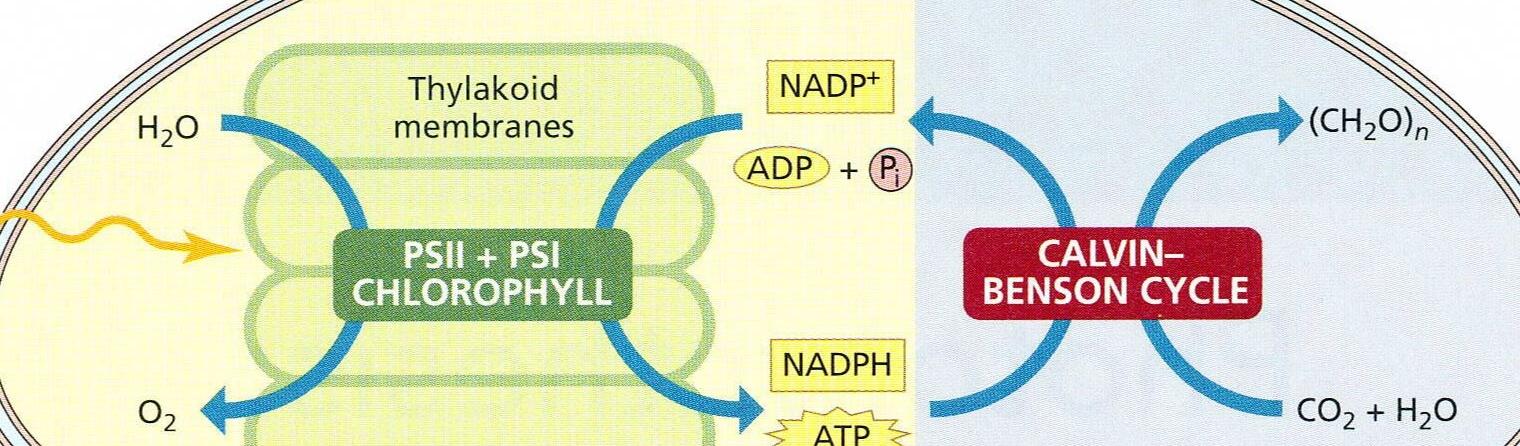
卡尔文循环 Calvin cycle
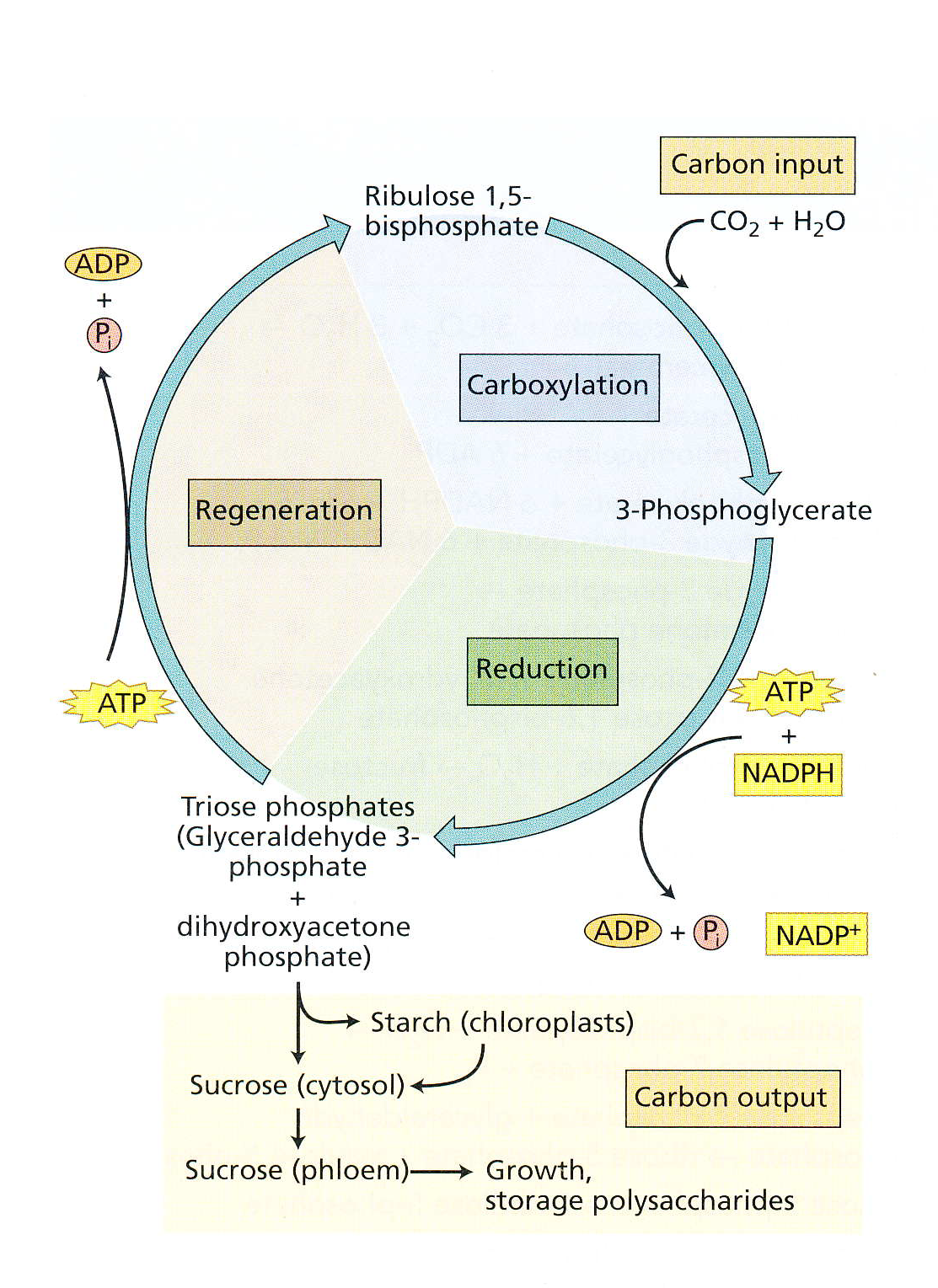
Calvin cycle
卡尔文循环又称 还原戊糖磷酸循环 ( reductive pentose phosphate cycle )。
如上图所示, 卡尔文循环分为三个阶段:羧化(caboxylation)、还原(reduction)、再生(regeneration)。
A. 羧化(carboxylation)
二氧化碳受体:核酮糖1,5 - 二磷酸 羧化,产生两分子 3 - 磷酸甘油酸;
B. 还原(reduction)
3 - 磷酸甘油酸还原,产生甘油醛 - 3 - 磷酸 和一种碳水化合物;
C. 再生(regeneration)
3 - 磷酸甘油酸再生,形成 核酮糖1,5 - 二磷酸。
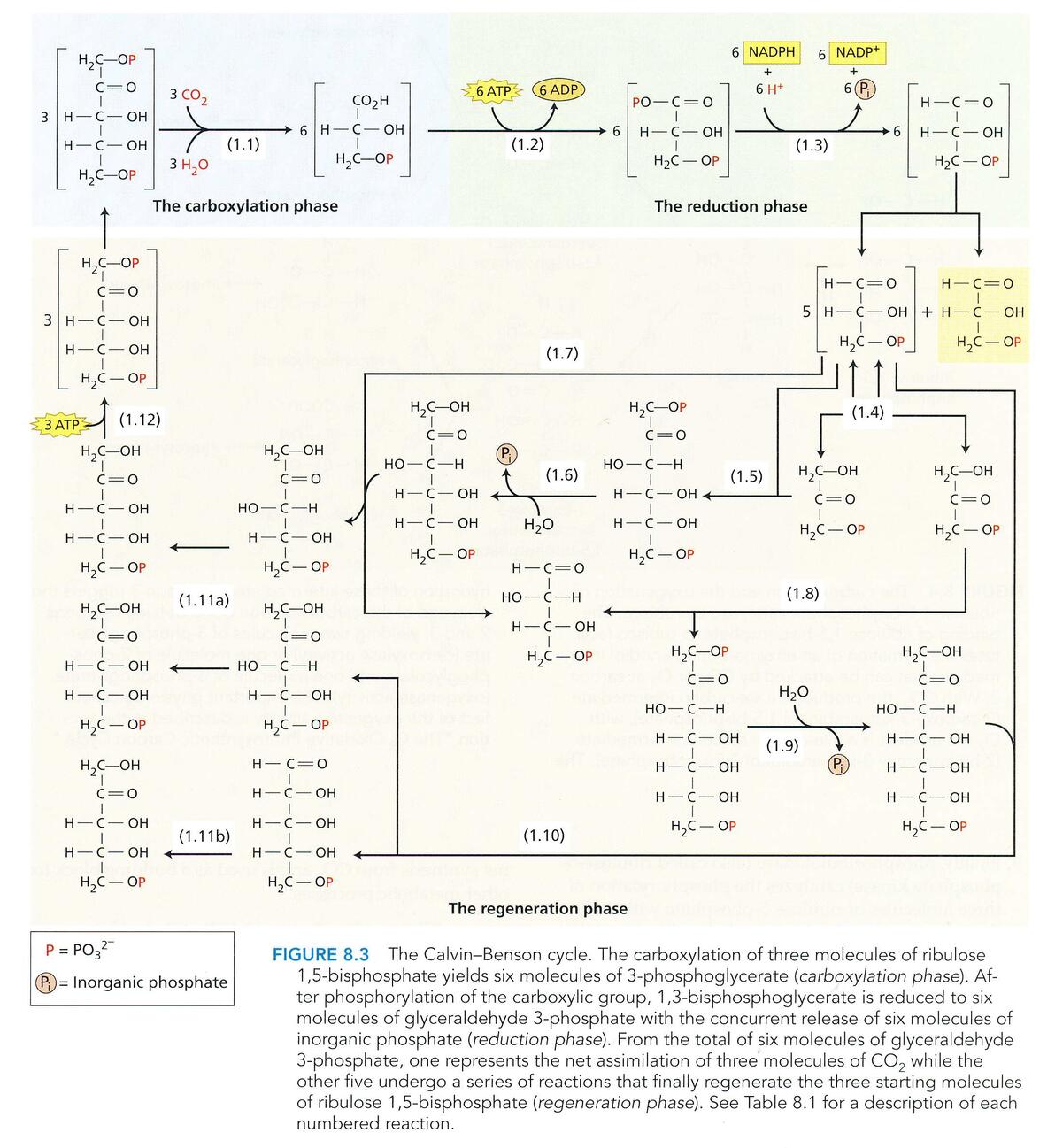
卡尔文循环详图
仔细看看上图,up想问一下大家,是不是很眼熟?
对,你没有看错,卡尔文循环中的还原模块里的确像极了前几章里提到的磷酸戊糖途径(PPP),就是原封不动的倒叙 ~ ^ _ ^ ~
这里要强调一下催化 核酮糖1,5 - 二磷酸 羧化的酶,Rubisco酶——
这个酶全称贼长:
核酮糖1,5 - 二磷酸 羧化/加氧酶 Ribulose-1,5-bisphosphate carboxylase/oxygenase
作为世界头号大酶,这个酶就是全球含量最高的蛋白质,没有之一,不接受反驳(笑),记得之前有人说,就算全球蛋白质加在一起,这个酶所占比重是能超过50%的。
Ellis RJ. 1979. Most abundant protein in the world. Trends in Biochemical Sciences 4: 241–244.
Losh JL, Young JN, Morel FMM. 2013. Rubisco is a small fraction of total protein in marine phytoplankton. New Phytologist 198: 52–58.
up下面就贴出Rubisco催化核酮糖-1,5-二磷酸羧化和加氧的详细化学反应,有兴趣的小伙伴可以深究一下,如果赶时间就跳过吧 ~ ^ _ ^ ~
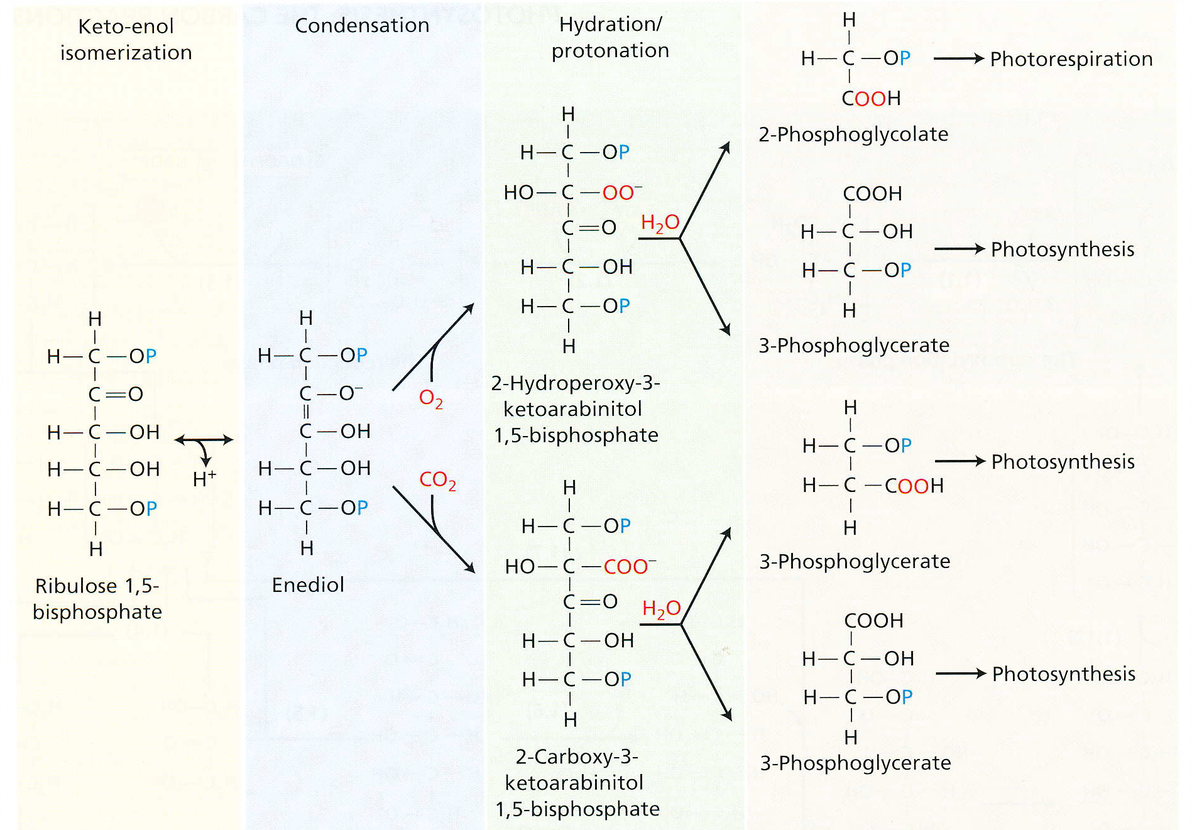
蓝条里,加氧在上,羧化在下,加氧最后形成的物质其一—— 2-磷酸乙醇酸(右侧第一个化学式)是用于光呼吸的。
卡尔文循环的继续进行,需要核酮糖-1,5-二磷酸的再生,以免该循环内的产物耗竭。
在该循环中,每固定一分子CO2,需要消耗 2 分子 NADPH 和 3 分子 ATP 才能合成碳水化合物。
让我们简单的计算一番,就能发现植物的精明之处:
1mol己糖生成CO2,氧化所释放的能量为 2804KJ,这也是合成己糖所需的最低能量;
6molCO2合成1mol己糖——果糖-6-磷酸 消耗为 3126 KJ,即:氧化12mol NADPH (12mol X 217 kJ/mol)和水解18mol ATP (18mol ATP X 29 kJ/mol),其热力学效率接近90%。
九成的功力啊!!!
应该还没有哪种人造发动机的转化效率能如此之高吧(虽然这个比喻可能不是很恰切)……生命如此可怕……
卡尔文循环的调控
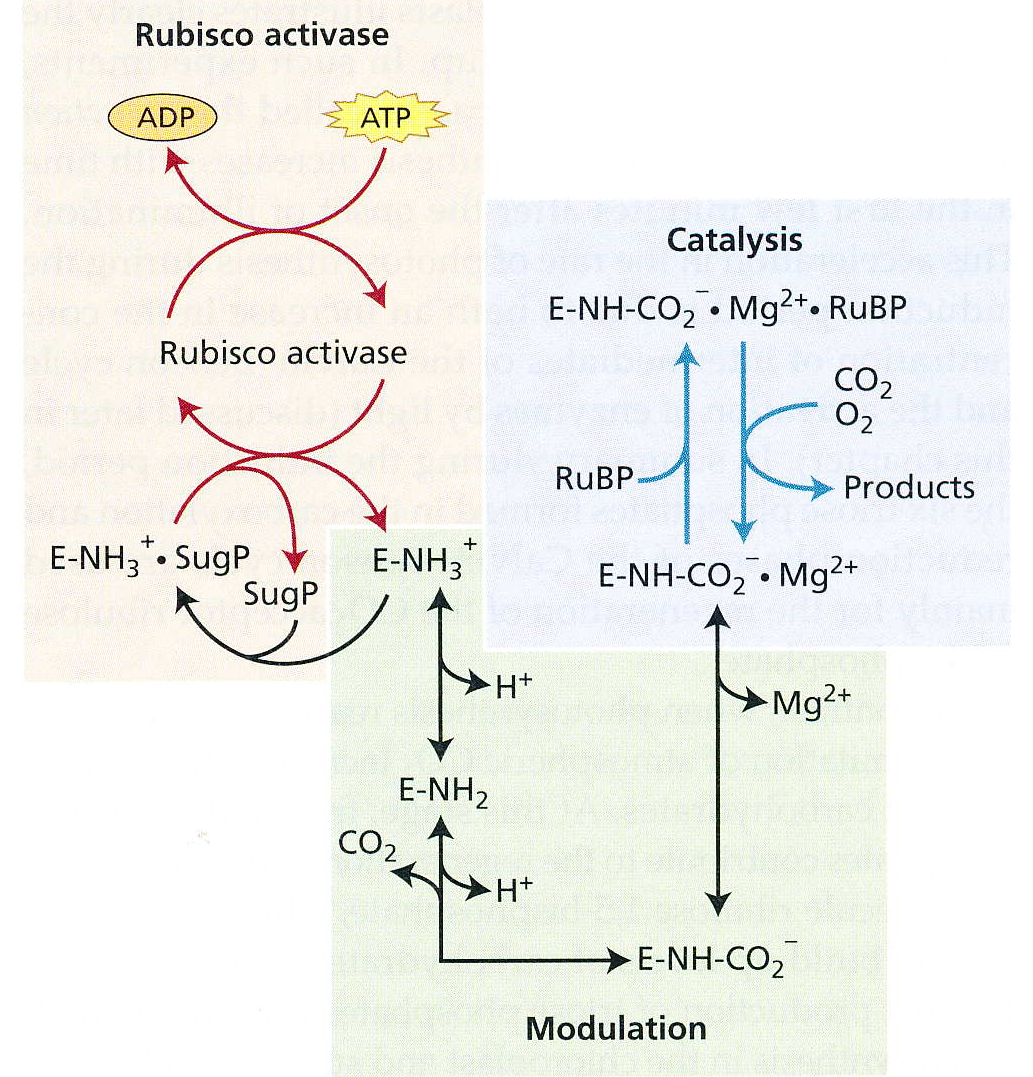
CO2作为Rubisco的激活剂(或者说,底物),能调节Rubisico的活性。
光控!!!光照能提高Rubisco的活性。
以下5个酶都是非常重要的调控开关:
Rubisco
果糖-1,6-二磷酸磷酸酶
景天庚酮糖-1,7-二磷酸磷酸酶
核酮糖-5-磷酸激酶
NADP-甘油醛-3-磷酸脱氢酶
其实除了Rubisco,光照还能通过 铁氧还蛋白-硫氧还蛋白系统来调节卡尔文循环中的其他四个酶。在此up并不想多写,有兴趣的小伙伴可以去翻一翻生理的课本,这儿应该超出了生化的范畴。
C2氧化光合碳循环
上图中,up提到,Rubisco既能给核酮糖-1,5-二磷酸 加氧,又能使核酮糖-1,5-二磷酸 羧化。而对于加氧的一部分,在光合作用活跃的叶片中,光依赖性的O2吸收和CO2的释放,构成了 光呼吸(photorespiration),这个过程会使卡尔文循环固定的CO2损失。
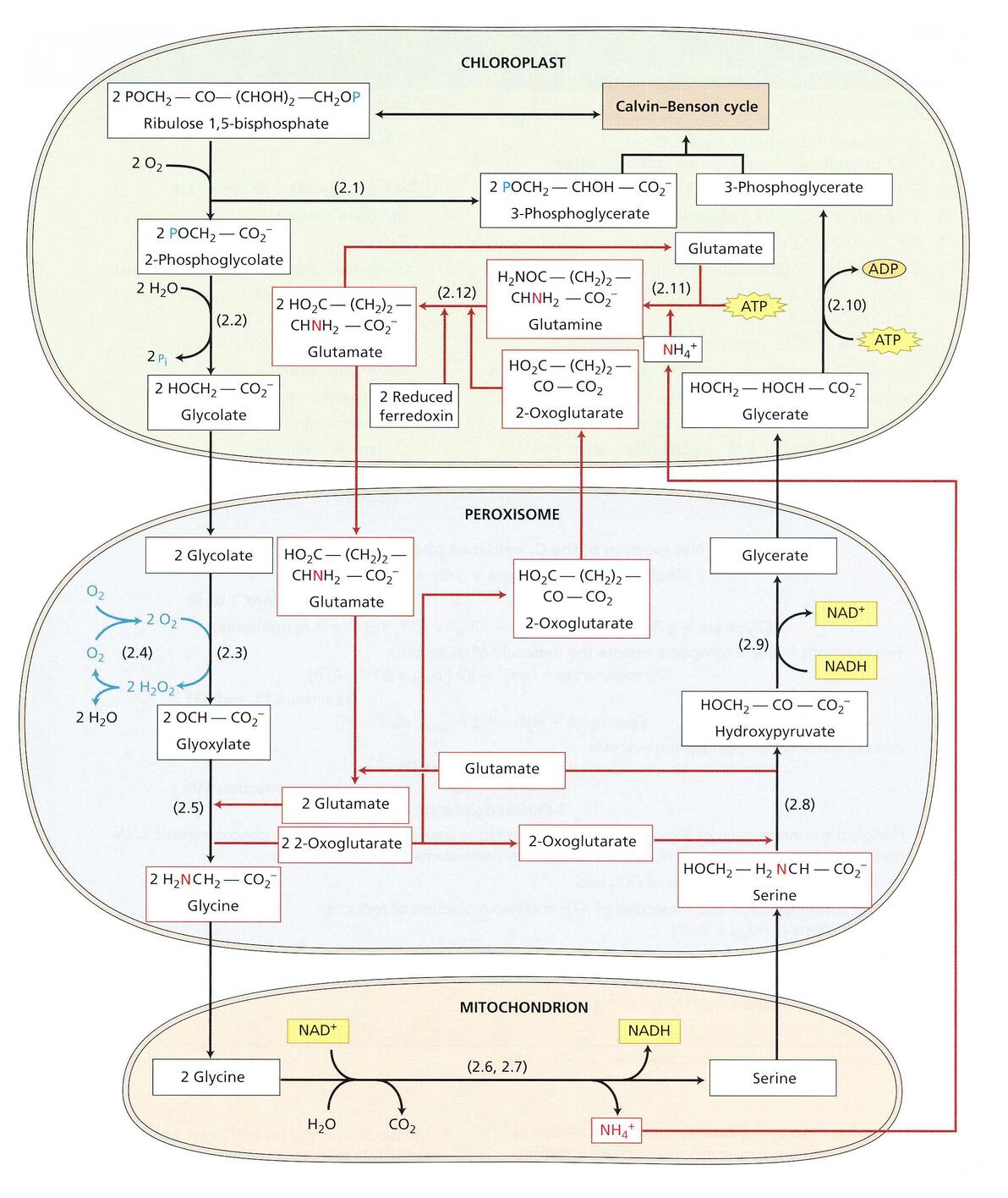
光呼吸循环的主要反应
而此处要说道说道的 C2氧化光合碳循环,使得光呼吸中损失的C得到了部分补偿(这个补救措施我是服的),下图是C2氧化光合碳循环的反应式:

C2氧化光合碳循环的反应方程式
光合CO2固定和光呼吸氧化是竞争反应,是一种此消彼长的状态。
总之,卡尔文循环的核酮糖-1,5-二磷酸的氧化所产生的2分子磷酸乙醇酸(4个C)转化为1分子的3-磷酸甘油酸(3个C)和一分子CO2。换言之,在核酮糖-1,5-二磷酸 的氧化过程里损失的75%的碳在C2氧化光合碳循环中,得到了弥补,返回到了卡尔文循环中。
相较于这种非光呼吸状况(低氧高CO2)来说,光呼吸状态下(高氧低CO2)固定同样多的CO2需要消耗更多的光能。
PS:高温,会使得Rubisco偏向于加氧(光呼吸),而不是羧化(光合)。总之:高温使得平衡偏向与C2氧化光合循环,而不是卡尔文循环。
科研界对光呼吸的生物学功能众口不一,有的说这玩艺儿和阑尾一样没什么卵用,有人坚持认为其形成了叶绿体中的一种辅助性保护机制用来减少ROS对叶绿体的伤害。简而言之,虽然不断有研究指出光呼吸的益处,但其至今并未完全被探明。
C4途径
刚刚上文我们也提到了,植物处于高温和水分胁迫的环境下,Rubisco的羧化酶活性会大大受到抑制,C4植物就针对该问题提出了一整套的解决方案(啪叽啪叽,Maize万岁!!赫鲁晓夫的胜利【滑稽】),这关键的浓缩碳机制,可以应对大气低CO2浓度造成的限制。
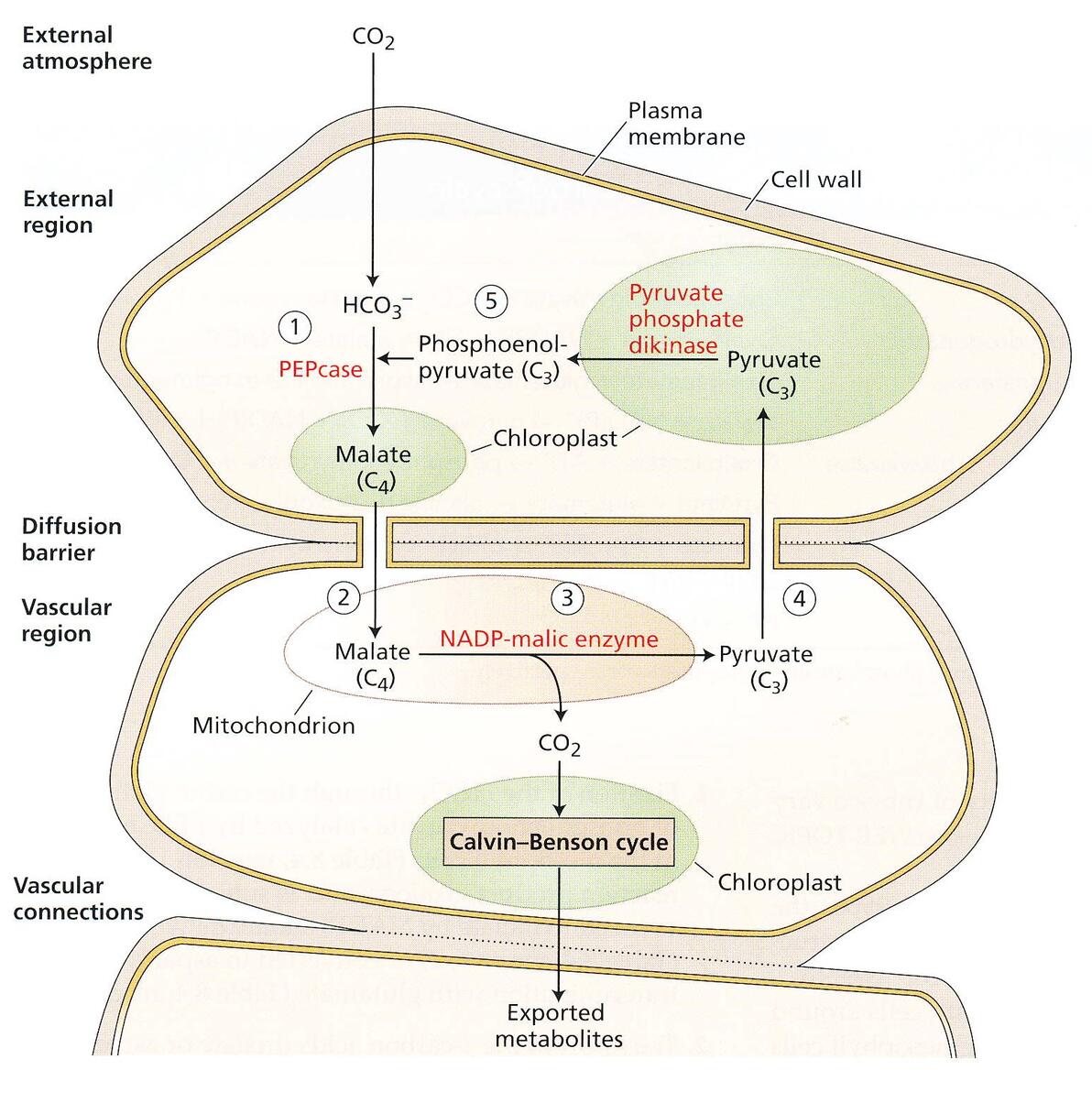
C4途径
C4途径光合作用,发生在一株C4植物的两种不同的维管组织中:
维管束鞘细胞(bundle sheath cell)和叶肉细胞(mesophyll cell)
叶肉细胞收集的CO2在PEPcase的作用下,和磷酸烯醇式丙酮酸(PEP)羧化形成苹果酸,而后运输到维管束鞘细胞,在NAD-苹果酸酶的催化下,释放出CO2,再进行碳反应。
这里普及一下洋文: kranz结构(花环结构)
实际上是在空间分区上实现了CO2富集。
PEPcase对CO2的捕获能力比Rubisco不知道高到哪里去了,所以C4作物(比如玉米)比C3作物(比如小麦)浓缩CO2的能力要高许多,从而带来了产量上的压倒性优势。
当然,在生命科学研究中,总会有打破常规的“异端”存在(笑),所以搞得现在的生命科学研究中的定义都不敢把话说死(怕打脸),自从发现了C4途径,人们习惯性认为这个过程不可能在一个细胞中出现,可是……可是……
Borszczowia aralocaspica 和 Bienertia cycloptera 就可以在一个细胞中完成这些活动。
Emmm……(陷入沉思,生命果然博大精深)
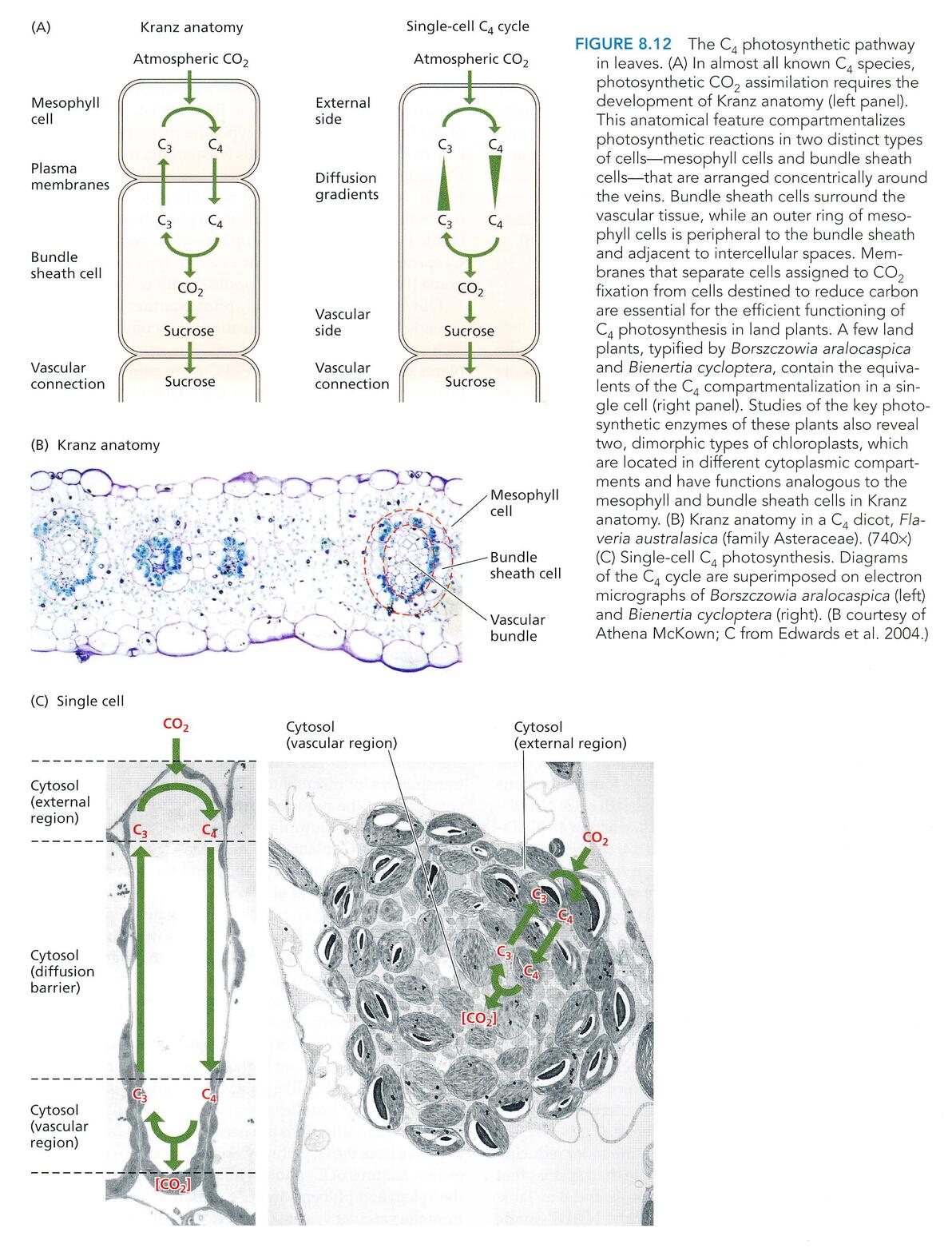
单细胞的C4光合作用
景天酸代谢(CAM)
在C4作物中,四碳酸的形成在一个区域(比如叶肉),脱羧和卡尔文循环又在另一个区域(比如维管束鞘细胞),两者在空间上是分离的,CAM植物中的四碳酸的形成在时间和空间上都是分离的。
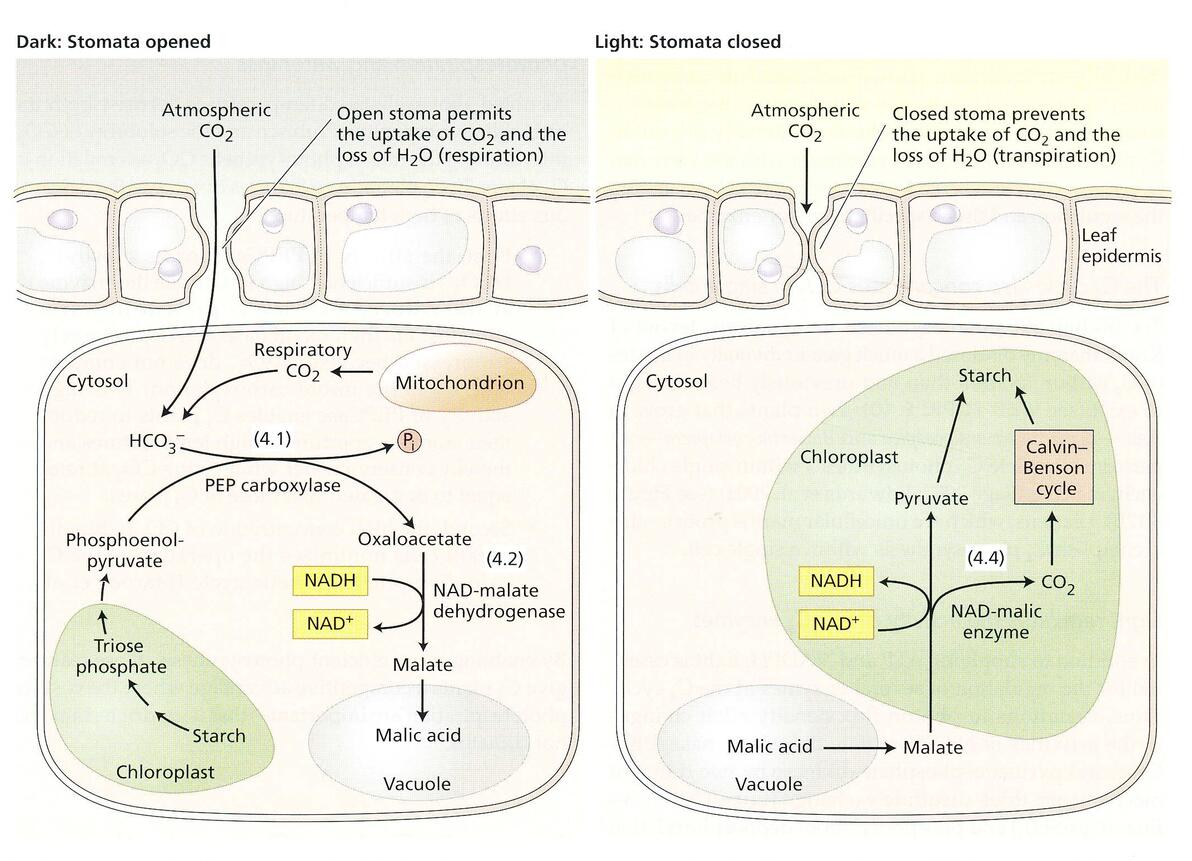
CAM途径
夜晚,报纸PEPcase捕捉CO2,形成草酰乙酸,而后在胞质的 NAD-苹果酸脱氢酶将草酰乙酸转化为苹果酸,储存于液泡中。
白天,贮存的苹果酸穿梭入叶绿体中,利用与C4植物类似的机制进行脱羧,然后释放的CO2用于卡尔文循环。
总之,恶劣的环境使得PEPcase和Rubisco催化的羧化反应在时间上分离,使得碳摄入和光合作用达到了最优化的状态。
总算打完了!!!!
哇的一声哭出来 …… > _ < ……
3000+啊~up更新小说都没有这么勤快过!up关于光合的笔记是英文版的,还是通篇翻译过来,今天更新晚了些,还望各位看官多多海涵。
之后就又回归手写的笔记了,下一章,是脂质的代谢~
尽请期待~
up溜啦~
