
细胞的凋亡,焦亡,坏死性凋亡共同构成了细胞从诞生到终结的复杂生命历程,本文将从简介,信号通路图,主要调控机制,相关产品等方面为您做详细介绍。
细胞凋亡(apoptosis)
1简介
细胞凋亡(Apoptosis)是一种程序性细胞死亡过程,是细胞在特定生理或病理条件下,通过一系列基因调控的有序过程,发生自主性细胞死亡,对维持生物体的正常发育、组织稳态和免疫调节等具有重要意义。
2信号通路图
3主要调控机制
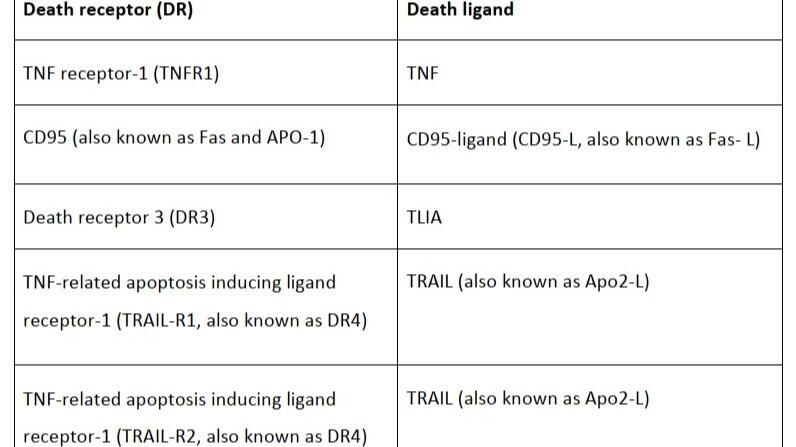
凋亡的起始取决于一系列半胱氨酸蛋白酶(Caspase)的激活。半胱氨酸蛋白酶(Caspase)分为两类:起始型半胱氨酸蛋白酶(Initiator Caspases)和效应型半胱氨酸蛋白酶(Executioner Caspases)。一旦检测到细胞损伤,起始型半胱氨酸蛋白酶(Caspase-8和Caspase-9)就会从无活性的前体形式被激活,并继续激活效应型半胱氨酸蛋白酶(Caspase-3,6和7)。效应型半胱氨酸蛋白酶的激活引发了一系列事件,从而导致核酸内切酶的激活,核蛋白和细胞骨架的破坏,蛋白质的交联和凋亡小体的形成等。
细胞凋亡过程在多细胞生物体内高度保守,并受基因控制。细胞凋亡可由细胞本身启动,当细胞通过一些细胞内传感器检测到自身损伤时会诱导细胞凋亡,这种机制被称为内源性途径。另外,免疫细胞与受损细胞之间的相互作用也会诱导细胞凋亡,这种机制被称为细胞凋亡的外源性途径。
(1)内源性信号通路
内源性途径,也称为线粒体凋亡途径,涉及作用于细胞内多个靶点的各种刺激。这种形式的细胞凋亡依赖于线粒体释放的因子,由应激信号激活。
细胞内的应激信号(如DNA损伤、氧化应激等)被细胞内的传感器识别,这些传感器包括p53、Bcl-2家族蛋白等。应激信号也可以通过激活促凋亡蛋白(如Bax、Bak、Bad)或抑制抗凋亡蛋白(如Bcl-2、Bcl-xL)诱发凋亡。
当细胞凋亡由应激性号诱发时,线粒体通透性会发生改变。促凋亡蛋白(包括细胞色素 c、Smac/Diablo 和 HtrA2/Omi)就能从线粒体泄漏到细胞质中,并激活细胞凋亡。
细胞色素 c 与 APAF1 单体的 WD 结构域结合,从而导致 APAF1 发生构象变化,多个 APAF1 聚集形成凋亡小体(apoptosome)。凋亡小体招募并激活 caspase 9,从而激活下游效应型半胱氨酸蛋白酶caspase 3等诱导凋亡。
Smac/Diablo 和 HtrA2/Omi 通过抑制凋亡抑制蛋白(IAPs)的表达从而协助启动细胞凋亡。
(2)外源性信号通路
外源性途径又称细胞凋亡的死亡受体途径,是由NK细胞或巨噬细胞产生死亡配体时启动,这些配体与靶细胞膜上的死亡受体(DR)结合后,通过激活caspase 8 诱导外源性途径。
死亡受体是TNF家族的成员,包括TNFR1、DR3、DR4等,每个死亡受体都有一个特定的死亡配体。
当死亡受体和对应的死亡配体(如YLIA、TNF和TRAIL等)结合后,招募procaspase 8到死亡诱导信号复合物(DISC)中并产生caspase 8。DISC 还包含FAS 相关死亡结构域 (FADD) 或 TNFR 相关死亡结构域 (TRADD) ,有助于促进procaspase 8 与 DISC 相互作用。
在 I 型细胞中,caspase 8 直接裂解和激活下游效应型caspases,从而直接启动细胞凋亡。在 II 型细胞中,IAPs 可抑制 Caspase 8 直接激活效应型Caspases,需要线粒体释放蛋白质(如Smac/Diablo 和 HtrA2/Omi )抑制IAPs,启动细胞凋亡。
活化的Caspase-8还可以激活Bax或Bak,促进线粒体膜通透性增加和细胞色素C释放,从而激活内源性途径,增强细胞凋亡信号。
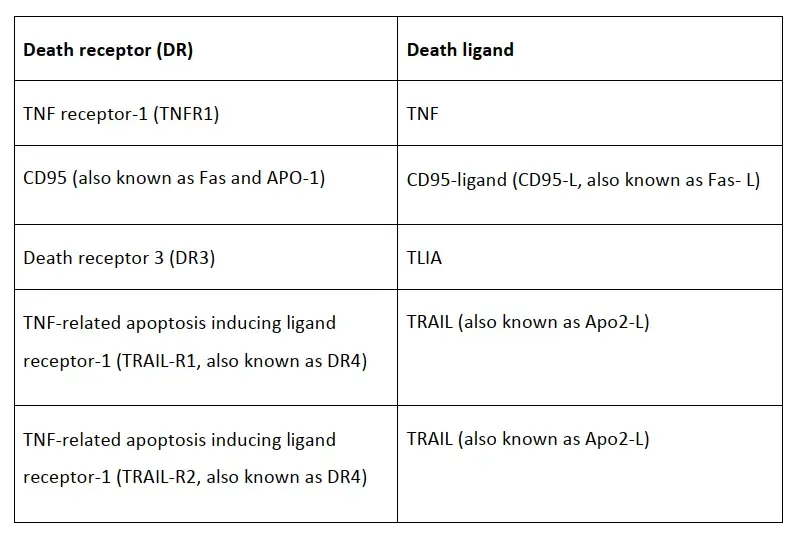
细胞死亡受体和对应的配体
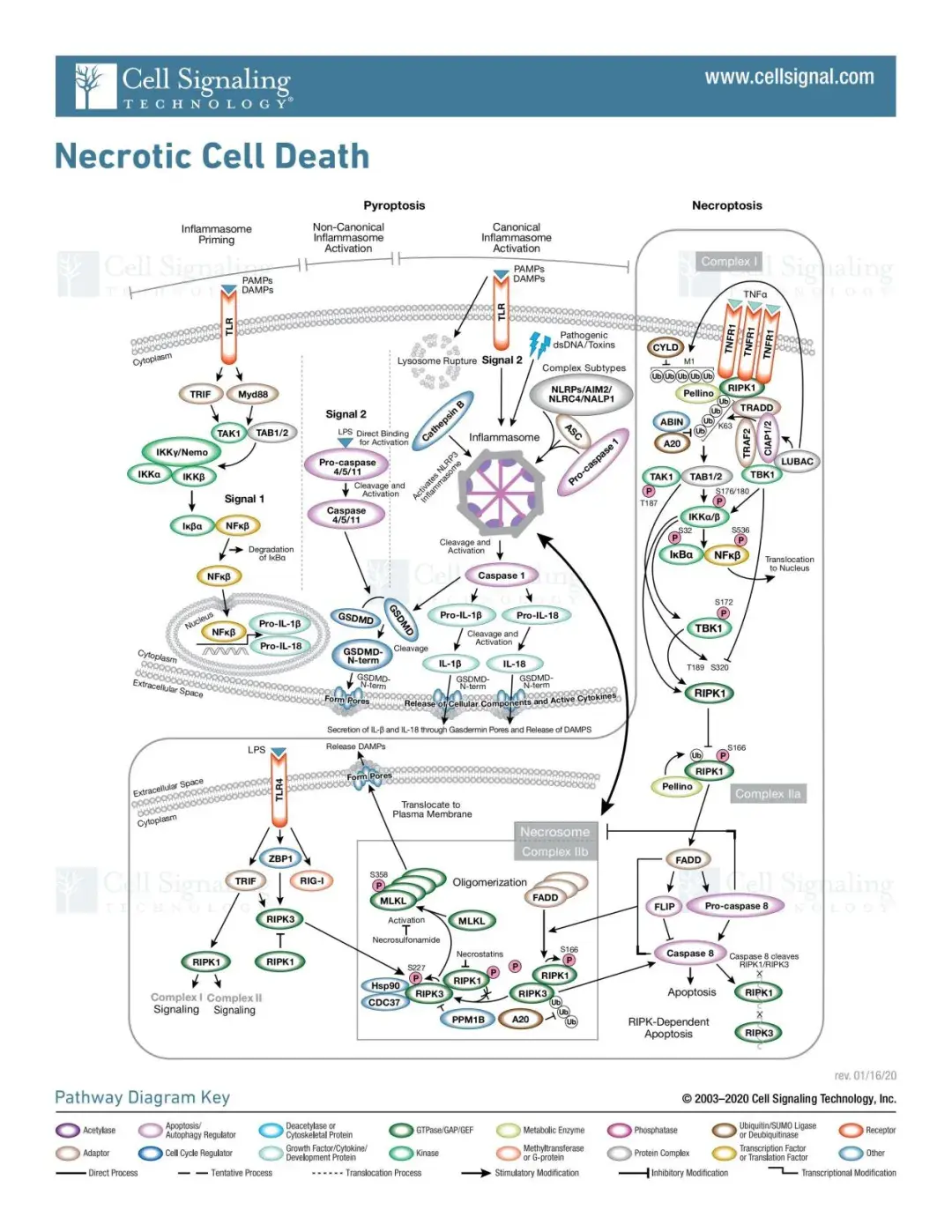
细胞焦亡(Pyroptosis)
1简介
细胞焦亡(Pyroptosis)是一种程序性的细胞死亡,其特征是细胞肿胀和渗透性裂解,导致细胞膜破裂和免疫刺激成分释放,这些成分在多种病理过程中发挥作用。
2信号通路图
3主要调控机制
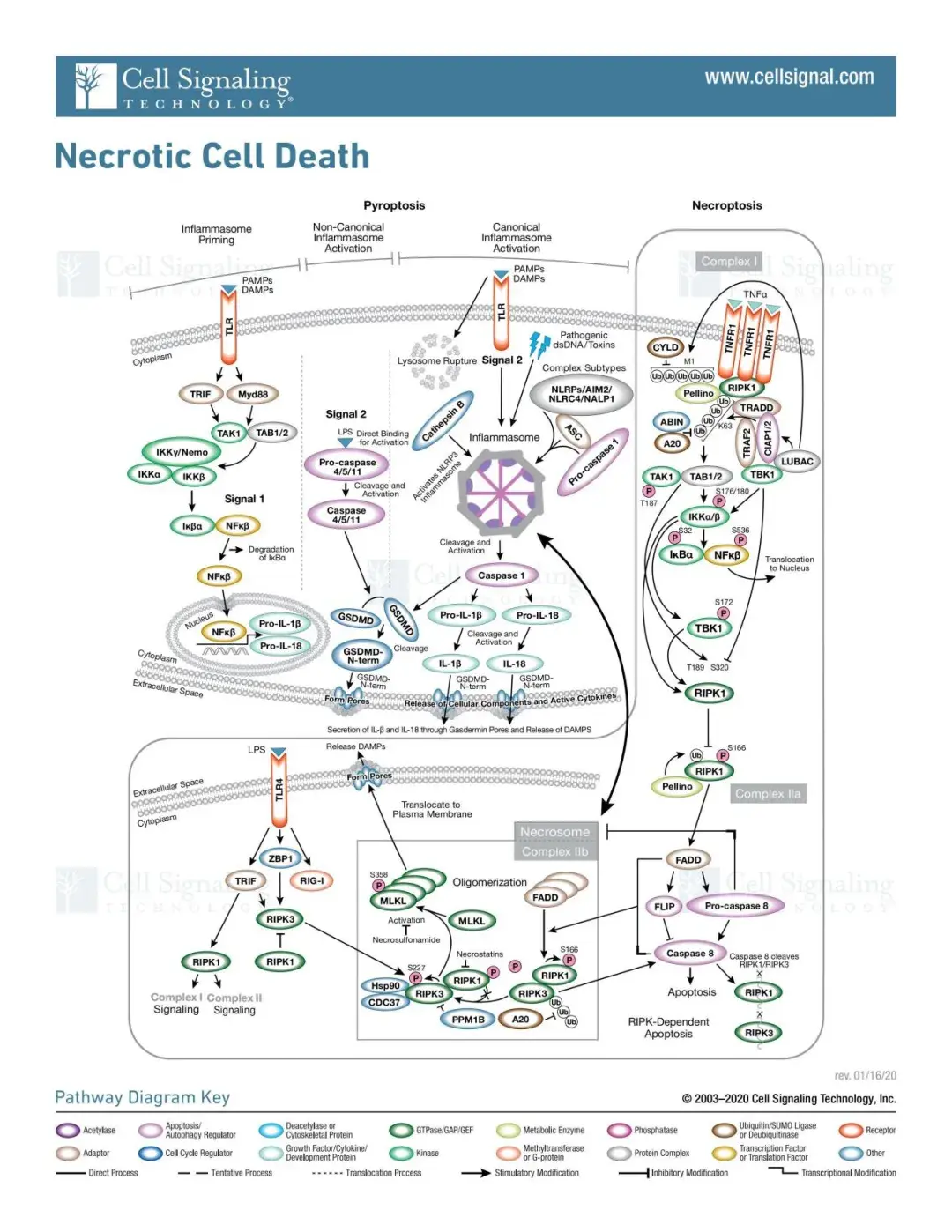
(1)经典途径
典型的细胞焦亡是由炎症小体的形成介导,伴随着 GSDMD 的裂解以及 IL-1β 和 IL-18 的释放。炎症小体的形成开始于细胞模式识别受体(PRRs,又称炎症小体传感器,如 NLRP1、NLRP3、NLRC4、AIM2等)识别病原体相关分子模式和危险相关分子模式(PAMPs 和 DAMPs),在细胞受到信号分子(例如细胞和病毒)刺激后,PRRs和pro-caspase-1 和 ASC 结合形成炎症小体。
炎症小体形成后,pro-caspase-1被激活并裂解为活性caspase-1。caspase-1 在 Asp275 位点裂解GSDMD,形成 22 kDa 的 C 端(C-GSDMD)和 31 kDa 的 N 端(NGSDMD)。N-GSDMD 可使细胞膜穿孔,形成内径约为 10-14 nm 的非选择性孔隙,导致细胞肿胀和细胞凋亡。
caspase-1 还会将 pro-IL-1β 和 pro-IL-18裂解为成熟的 IL-1β 和 IL-18,并通过 GSDMD 形成的孔释放出来,从而导致细胞焦亡。
(2)非经典途径
在非经典焦亡途径中,人类 Caspase-4/5(小鼠同源物 Caspase-11)的上游感受器复合体缺失,这些Caspases的N端CARD结构域可直接与细胞内脂多糖(LPS)结合而被激活。
活化的 Caspase-4/5/11裂解GSDMD,释放N-GSDMD,形成细胞膜孔。然而,caspase-4/5/11不能裂解pro-IL-1β/pro-IL-18,但在某些细胞中,它们能够通过NLRP3/caspase-1途径介导IL-1β/IL-18的成熟和分泌。
(3)Caspase-3/8-mediated pathway
化疗药物可以诱导 caspase-3 激活,并裂解Gasdermin E(GSDME),释放N-GSDME 末端,从而引起肿瘤细胞的焦亡。
此外,在小鼠巨噬细胞中发现,耶尔森菌感染期间,效应蛋白 YopJ 可抑制 TAK1的表达,并诱导Caspase-8 相关的 GSDMD 裂解,进而引起细胞焦亡。
其他发现包括PD-L1 在乳腺癌细胞中能将 TNF 介导的凋亡转化为焦亡。在缺氧条件下,p-Stat3 促进 PD-L1 的核转位,并共同增强 GSDMC 的转录。
在 TNF-α 的刺激下,Caspase-8 会特异性地裂解 GSDMC,形成 N-GSDMC,并在细胞膜上成孔诱导细胞焦亡。巨噬细胞来源的 TNF-α 诱导的肿瘤焦亡需要核 PD-L1、caspase-8 和 GSDMC。此外,抗生素化疗药物也会引起caspase-8/ GSDMC 介导的乳腺癌细胞焦亡。
(4)颗粒酶介导的途径
细胞毒性淋巴细胞(如自然杀伤细胞和细胞毒性T细胞)通过释放颗粒酶A(GzmA)和颗粒酶B(GzmB)通过穿孔素进入靶细胞,诱导焦亡。
GzmA可水解GSDMB,并激活GSDME诱导细胞焦亡。GzmB可快速激活靶细胞中的 caspase-3,然后激活 caspase-3/GSDME 介导的焦亡途径,引起广泛的细胞焦亡。GzmB 也可直接裂解 GSDME 并诱导细胞焦亡,从而进一步激活抗肿瘤免疫反应并抑制肿瘤生长。
细胞坏死性凋亡(Necroptosis)
1简介
Necroptosis(坏死性凋亡)是一种程序性细胞坏死方式,特征是细胞肿胀、细胞膜完整性丧失以及细胞内容物的释放,这些特征与细胞坏死相似,但受到特定信号通路的精细调控。
2信号通路图
3主要调控机制
(1)经典途径
细胞坏死性凋亡是在TNFR1被 TNFα 激活后发生的。其他细胞受体的激活会引发坏死,这些受体包括死亡受体(例如Fas/FasL),Toll 样受体(TLR4 和 TLR3)和细胞膜核酸感受器器(例如 RIG-I 和 STING),它们可诱导IFN-I和 TNFα 的产生,从而促进细胞坏死。
这些途径大多会触发NFκB 依赖的促炎症和促生存信号。然而,微生物或药物对Caspase-8 的额外抑制也会触发坏死途径。
在上述受体的下游,活性 RIPK1 被招募到一个包括 FADD、caspase-8 和 caspase-10 在内的复合物中。在缺乏 caspase-8 活性的情况下,RIPK1 会招募并磷酸化RIPK3,形成坏死小体。RIPK1/RIPK3 复合物招募并磷酸化MLKL,从而形成坏死体。
MLKL进一步损伤细胞完整性:(1)MLKL 在质膜上构成一个平台,用于打开钙离子或钠离子通道,从而使离子流入、细胞肿胀和破裂(2)MLKL直接与质膜上的PIPs相互作用,在质膜上形成孔隙。
(2)其他非经典途径
CYLD是一种去泛素化酶,它可以特异性的去泛素化TRAF2 和 RIPK1,从而形成坏死小体,启动坏死性凋亡;线粒体蛋白 Smac 通过触发蛋白酶体降解 cIAP1/2 和 XIAP,解除他们对坏死性凋亡的抑制作用,PIPK1和PIPK3形成坏死小体,促进坏死性凋亡;此外DNA损伤,氧化应激,缺氧,病毒感染等途径也会激活PIPK1和PIPK3从而引发坏死性凋亡。
