
高清版信号通路图可联系销售索取
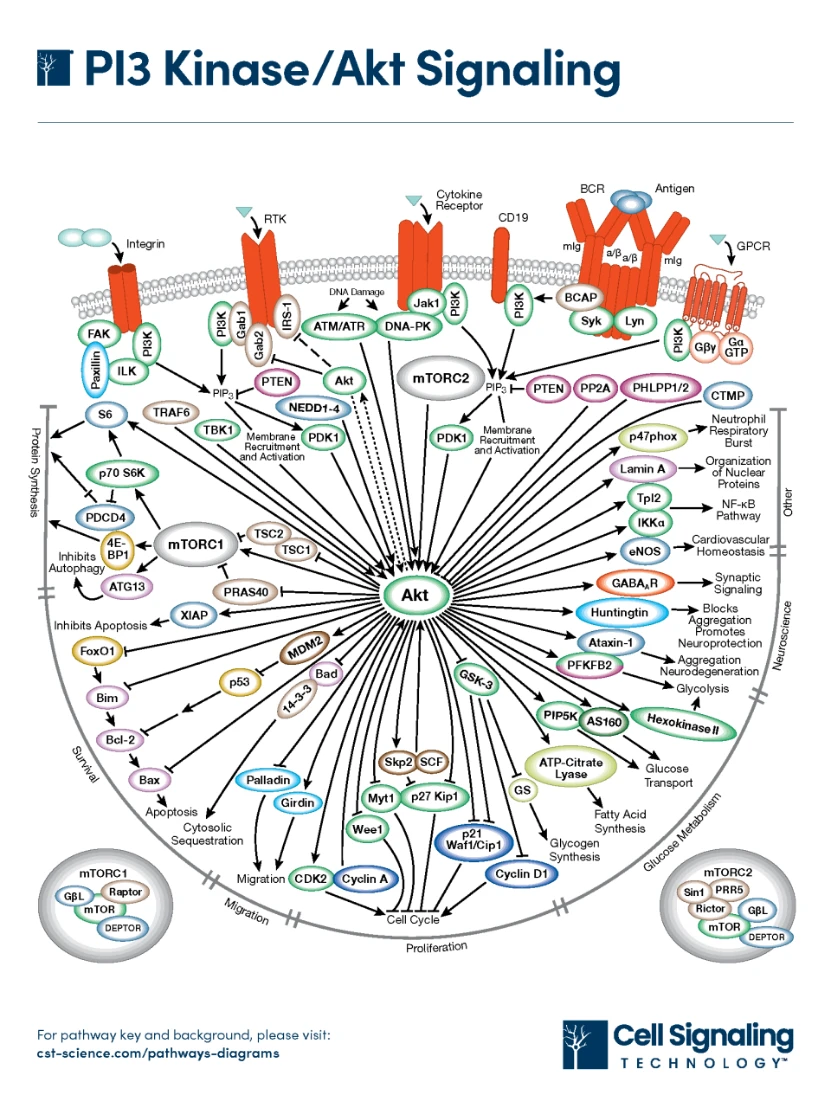
PI3K/AKT/mTOR(PAM)信号通路是真核细胞中高度保守的信号转导网络,作用于促进细胞存活、生长和细胞周期进程。PAM轴上生长因子向转录因子的信号传导受到与其他多条信号通路的多重交叉相互作用的严格调控,信号转导失调可能导致疾病发生发展。1
核心分子
PI3K
PI3K是细胞内脂质激酶家族的成员之一,主要由p55和p85调节亚基以及p110催化亚基构成2。根据亚基和底物的不同,PI3K主要分为三类,即 I 类、II 类和 III 类。
I 类 PI3K 是由 p110 催化亚基和 p85 调节亚基组成的异二聚体,有IA类(PI3Kα 、PI3Kβ和PI3Kδ )和IB类(PI3Kγ )四种亚型3 ,通过激活下游酪氨酸激酶(例如 G 蛋白偶联受体 (GPCR) 和小单体 GTP 酶)发挥作用。此外,p85 亚基还可以传递各种细胞信号,为信号整合和下游蛋白质的激活奠定关键基础。4
II 类 PI3K 包括 C2α、C2β 和 C2γ 催化亚型,缺乏调节亚基,因此可以作为单体被激活。在哺乳动物中,已鉴定出三种 II 类 PI3K 亚型,其中 PI3KC2α 和 PI3KC2β 广泛表达,而 PI3KC2γ 主要在肝脏中表达。4
III 类 PI3K VPS34(也称为 PIK3C3)非常独特,它通过结合由调节亚基和催化亚基组成的蛋白质复合物,在调节自噬和巨噬细胞吞噬中发挥重要作用。因此,异二聚体 III 类 PI3K 也能调节自噬。4
AKT
AKT属于丝氨酸/苏氨酸激酶家族成员,由普列克底物蛋白同源(PH)结构域、中间激酶结构域和调节性羧基末端组成,主要包括AKT1、AKT2和AKT3亚型。Akt1 和 Akt2 在许多组织中富集,例如胰腺组织,而 Akt3 主要在脑中表达,并且其表达受组织分布的限制。4
上游激活
PI3K/Akt信号通路上游的激活与多种因素相关,包括RTK家族、Toll样受体(TLR)和B细胞抗原受体(BCR)。4
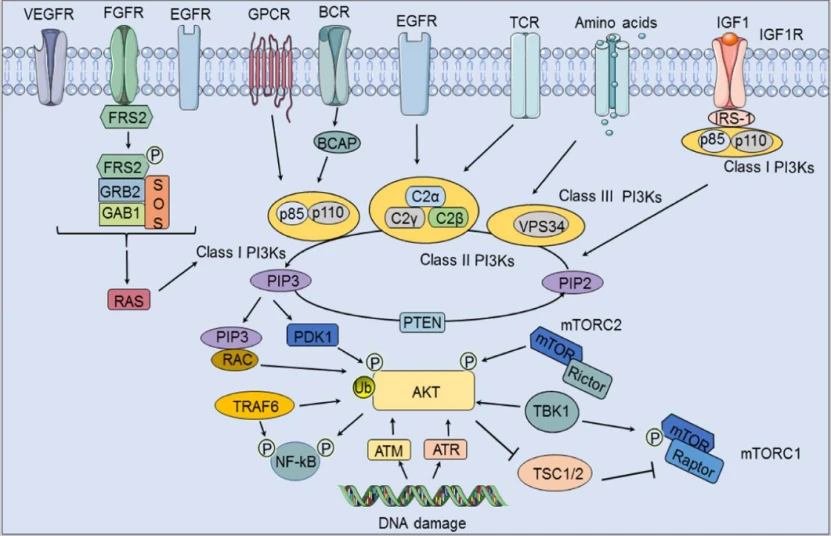
一方面,与特定 RTK(EGFR、VEGFR 和 FGFR)结合的配体可通过 RAS 激活 I 类 PI3K;另一方面,I 类 PI3K 可被 BCR 通过 B 细胞衔接子激活,也可被 GPCR 激活。FGFR 底物 FRS2 与 GRB2、SOS 和 GAB1 结合磷酸化,从而激活 I 类 PI3K。
II 类 PI3K 除可被 EGFR 激活外,还可被 TCR 激活。
III 类 PI3K 由氨基酸激活,而总活化 PI3K 会磷酸化 PIP2 肌醇头的第三个碳原子,并将其转化为 PIP3,从而通过 PDK1 和 RAC 激活 AKT,而该转化过程可被 PTEN 抑制。
此外,IGF-1与IGF1R结合可募集IRS-1和I类PI3K,进而参与PIP2向PIP3的转化。mTORC2可以通过影响Akt的磷酸化来影响Akt的活性,进而通过TSC1/2影响下游的mTORC1,而Akt和mTORC1均可被TBK1激活。TRAF6可以通过影响Akt的泛素化来影响Akt的活性。此外,DNA损伤可以通过ATM和ATR影响Akt。4
RTK
RTK 是一个具有内在磷酸酪氨酸激酶活性的跨膜蛋白家族,主要包括表皮生长因子受体 (EGFR)、血管内皮生长因子受体 (VEGFR) 和成纤维细胞生长因子受体 (FGFR)。同源生长因子、细胞因子和激素等配体通过激活 RTK 来激活 PI3K 信号通路。此外,I 类 PI3K 的 p85 亚基与磷酸化的 RTK 结合,导致 PI3K 催化结构域 (p110) 发生构象变化。4
TLR
TLR 在免疫细胞和非免疫细胞中表达,可以识别细菌脂多糖(由 TLR4 识别)和单链或双链 RNA(由 TLR3 识别)中保守的微生物基序,还能区分并结合内源性配体,从而激活信号通路。
BCRs
BCR激活的信号通路对B细胞的发育、活化和分化至关重要。其中,PI3K/Akt通路尤为重要。在B细胞中,I类PI3K由BCR通过B细胞受体相关蛋白(BCAP)激活,这是PIP3生成和Akt激活的重要步骤。BCR和胞质衔接子对PI3K/Akt信号通路的激活有显著影响,当B细胞缺乏BCR时,Akt不会被激活。
GPCR(G蛋白偶联受体)
GPCR 是最大的细胞表面蛋白家族,在细胞信号转导中发挥重要作用,也是 PI3K/Akt 信号通路的共同靶点。GPCR通过异三聚体 G 蛋白传递信号,并通过与直接结合在 GPCR 上并参与信号网络调控的各种小 G 蛋白相互作用来调控下游效应通路。
PTEN
PTEN 是一种肿瘤抑制基因,对维持正常的生理活动至关重要。作为一种脂质磷酸酶,PTEN 负向调控 PI3K 信号通路,并将 PIP3 转化为 PIP2。当PTEN 发生突变或参与其他形式的失活时,PI3K 效应分子(尤其是 Akt)会在没有任何外源致癌刺激的情况下被激活。
下游主要效应分子
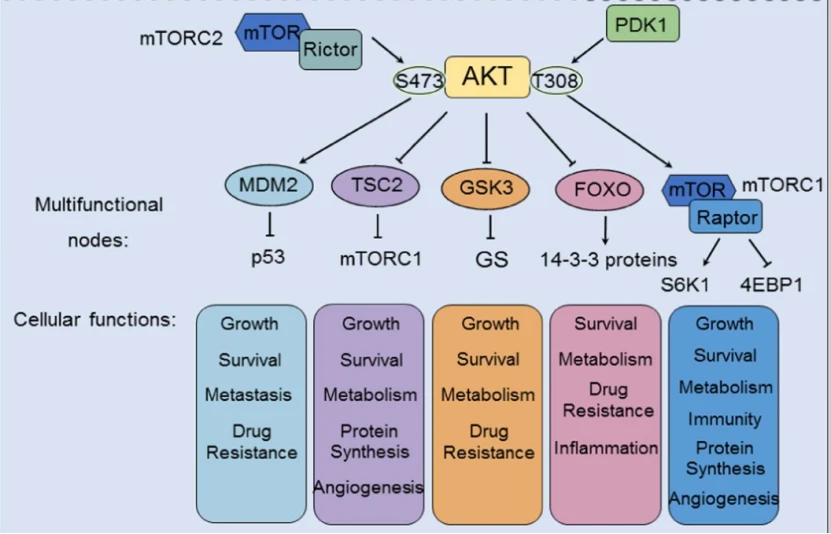
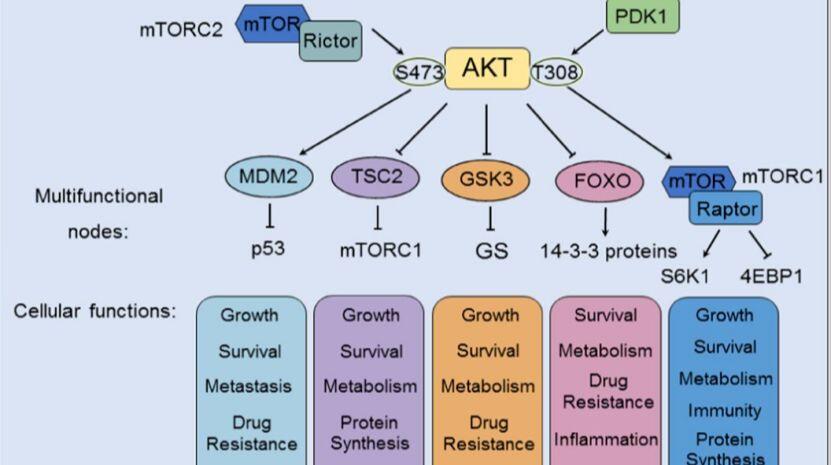
Akt信号的激活可以促进(箭头)或抑制(阻断箭头)下游效应分子的磷酸化。Akt的下游调控参与了许多细胞过程,包括肿瘤生长、肿瘤存活、肿瘤细胞增殖、癌症免疫、癌症代谢和癌症血管生成。4
mTOR
mTOR 参与调节肿瘤的生长、存活、代谢和免疫,通常组装成复合物,例如 mTORC1 和 mTOR 复合物 2(mTORC2),在许多生物过程中发挥关键作用,其抑制剂在临床癌症治疗中具有巨大的潜力。4
GSK3
GSK3是首个被报道的Akt底物,目前已鉴定出两种GSK3亚型:GSH3α和GSK3β。在 PI3K 介导的信号转导后,Akt 诱导 N 端调控结构域中的 Ser21 (α) 或 Ser9 (β) 磷酸化后,GSK3(GSH3α 和 GSK3β)失活并被蛋白酶体降解。GSK3的表达影响癌症中的各种生化过程,除了肿瘤生长之外,GSK3 还通过磷酸化和抑制代谢酶(例如其底物糖原合酶 (GS))参与肿瘤代谢。4
FOXOs
FOXOs是含FOX转录因子(TF)超家族的一个亚组。FOXO TFs 包含四个 Akt 的直接下游靶点:FOXO1、FOXO3、FOXO4 和 FOXO6。PI3K/Akt信号通路通过磷酸化FOXOs促进其与14-3-3蛋白结合,进而遮蔽NLS(阻断核转位并加速核输出),最终驱动泛素-蛋白酶体途径(UPP)依赖性降解,该机制是胰岛素/PI3K/Akt通路调控FOXO水平的核心方式。Akt耗竭的主要表型由FOXO介导的转录驱动,FOXOs是Akt信号在多种生物反应中的核心下游效应因子。
TSC2
生长因子(如IGF-1)通过进化保守的PI3K-Akt信号通路促进细胞生长,其核心机制依赖于Akt介导的TSC2(tuberin)磷酸化。TSC1/TSC2复合物作为由肿瘤抑制基因编码的负调控因子(其突变导致结节性硬化症),通过TSC2的GTP酶激活蛋白(GAP)结构域将Rheb-GTP(mTORC1的强效激活因子)转化为Rheb-GDP,从而抑制mTORC1活性;而Akt对TSC2的磷酸化逆转了这一抑制作用,解除TSC2对Rheb和mTORC1的抑制,最终激活mTORC1信号轴驱动细胞生长。
MDM2
PI3K/Akt信号通路的激活通过磷酸化MDM2的丝氨酸残基促进其核转位,进而增强该癌基因与抑癌蛋白p53的结合,抑制p53的转录活性并诱导其泛素化降解;此过程揭示了MDM2通过拮抗p53发挥促肿瘤作用的分子机制(其mRNA水平本身受p53调控)。PTEN对PI3K/Akt通路的抑制可保护p53免受MDM2介导的降解,且p53表达水平与DNA损伤应答呈正相关,共同确立了PI3K/Akt-MDM2-p53轴在细胞周期阻滞与凋亡调控中的核心地位。
